

The molecular genealogy of sequential overlapping inversions implies both homologous chromosomes of a heterokaryotype in an inversion origin
- PMID: 31740730
- PMCID: PMC6861252
- DOI: 10.1038/s41598-019-53582-8
The molecular genealogy of sequential overlapping inversions implies both homologous chromosomes of a heterokaryotype in an inversion origin
Abstract
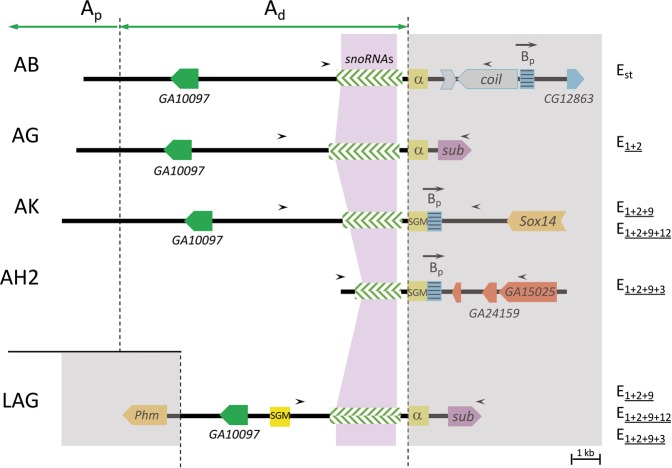
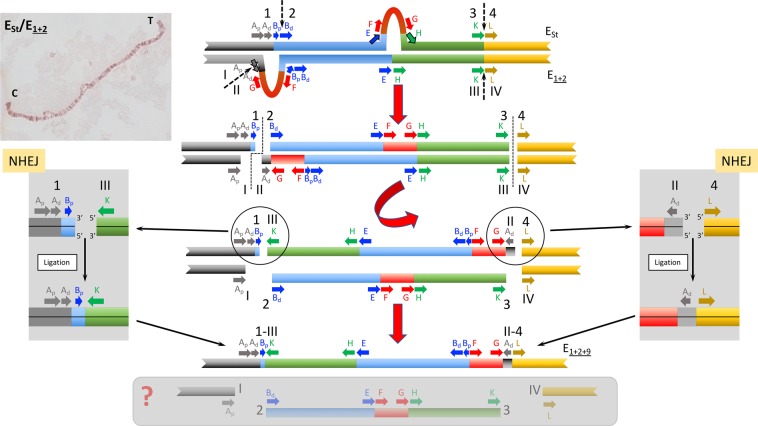
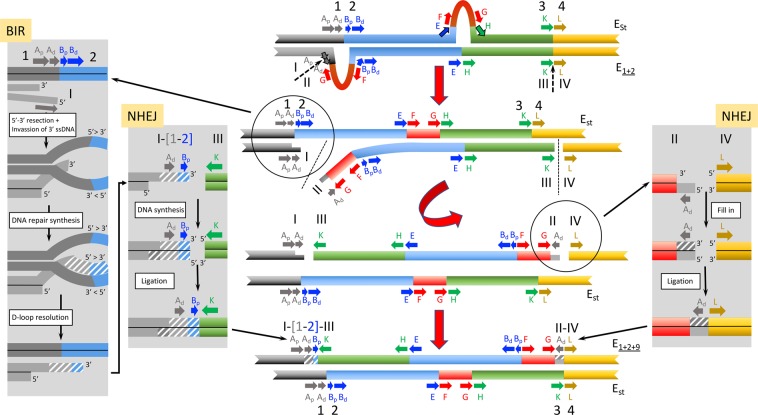
Cytological and molecular studies have revealed that inversion chromosomal polymorphism is widespread across taxa and that inversions are among the most common structural changes fixed between species. Two major mechanisms have been proposed for the origin of inversions considering that breaks occur at either repetitive or non-homologous sequences. While inversions originating through the first mechanism might have a multiple origin, those originating through the latter mechanism would have a unique origin. Variation at regions flanking inversion breakpoints can be informative on the origin and history of inversions given the reduced recombination in heterokaryotypes. Here, we have analyzed nucleotide variation at a fragment flanking the most centromere-proximal shared breakpoint of several sequential overlapping inversions of the E chromosome of Drosophila subobscura -inversions E1, E2, E9 and E3. The molecular genealogy inferred from variation at this shared fragment does not exhibit the branching pattern expected according to the sequential origin of inversions. The detected discordance between the molecular and cytological genealogies has led us to consider a novel possibility for the origin of an inversion, and more specifically that one of these inversions originated on a heterokaryotype for chromosomal arrangements. Based on this premise, we propose three new models for inversions origin.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Krimbas, C. B. & Powell, J. R. Drosophila inversion polymorphism. (CRC Press, 1992).
-
- Coluzzi M, Petrarca V, Di Deco MA. Chromosomal inversion intergradation and incipient speciation in Anopheles gambiae. Bolletino di Zool. 1985;52:45–63. doi: 10.1080/11250008509440343. - DOI
-
- Gupta JP, Kumar A. Cytogenetics of Zaprionus indianus Gupta (Diptera: Drosophilidae): Nucleolar organizer regions, mitotic and polytene chromosomes and inversion polymorphism. Genetica. 1987;74:19–25. doi: 10.1007/BF00055090. - DOI
