

Structural basis of amino acid surveillance by higher-order tRNA-mRNA interactions
- PMID: 31740854
- PMCID: PMC6899168
- DOI: 10.1038/s41594-019-0326-7
Structural basis of amino acid surveillance by higher-order tRNA-mRNA interactions
Abstract
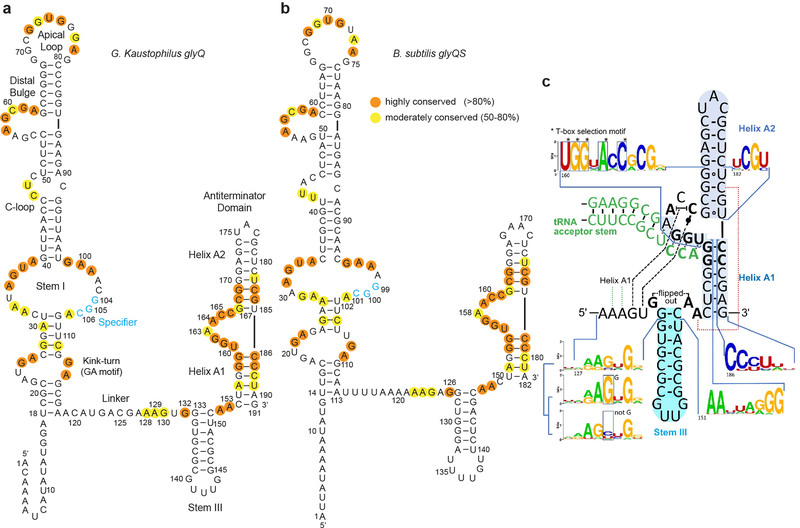
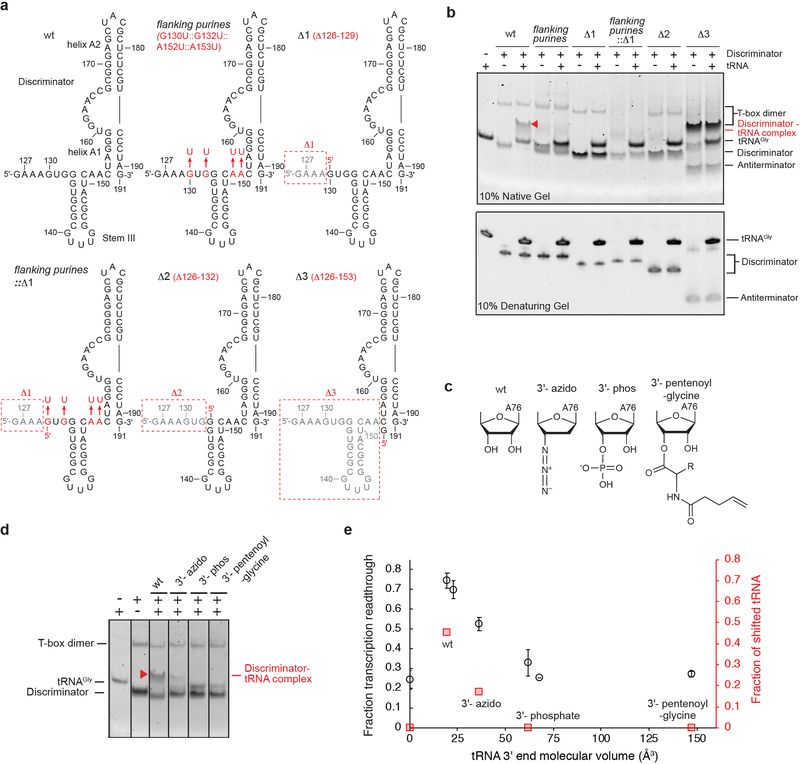
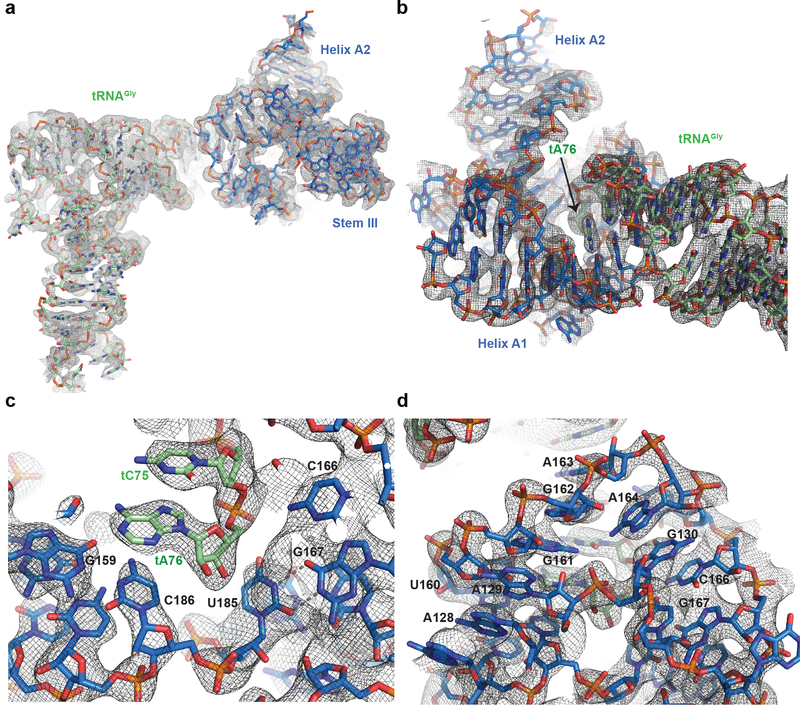
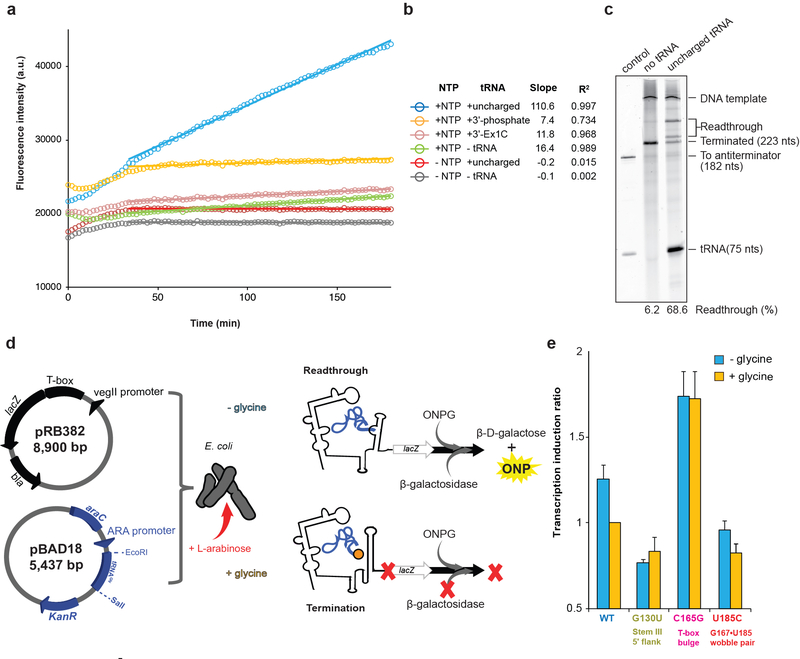
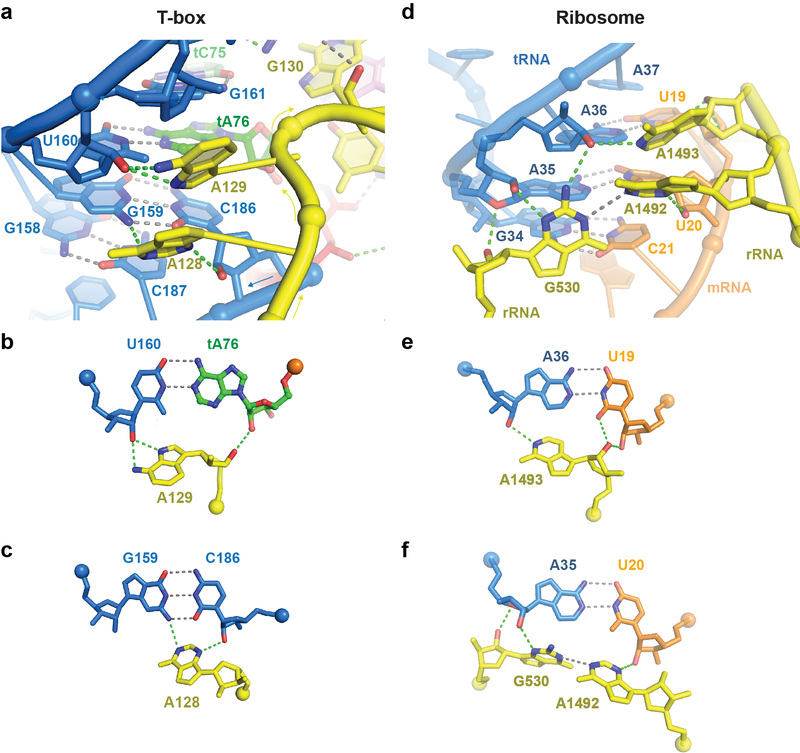
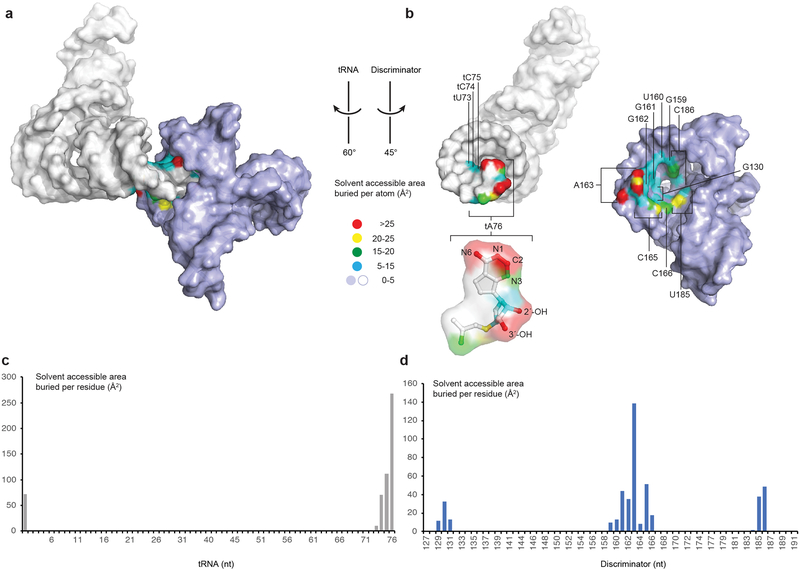
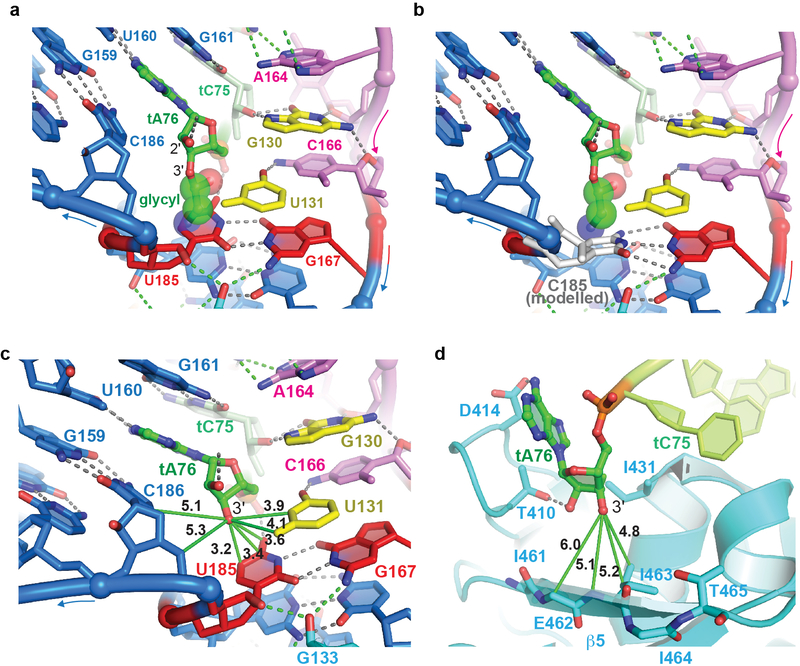
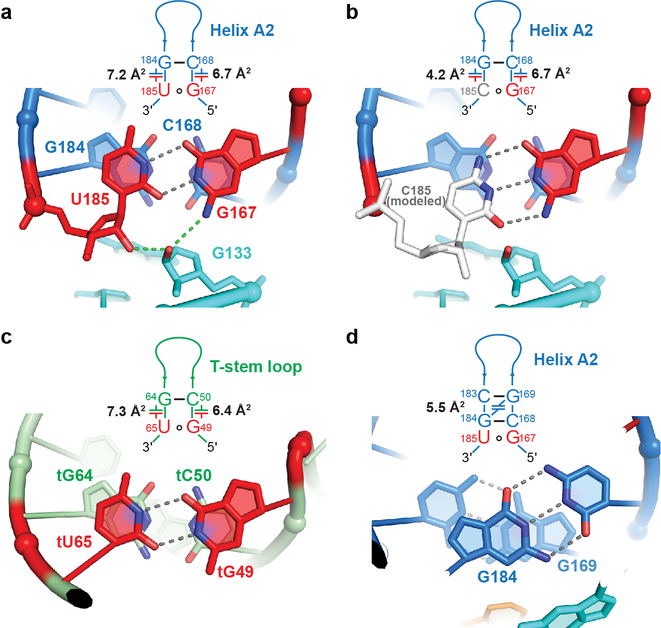
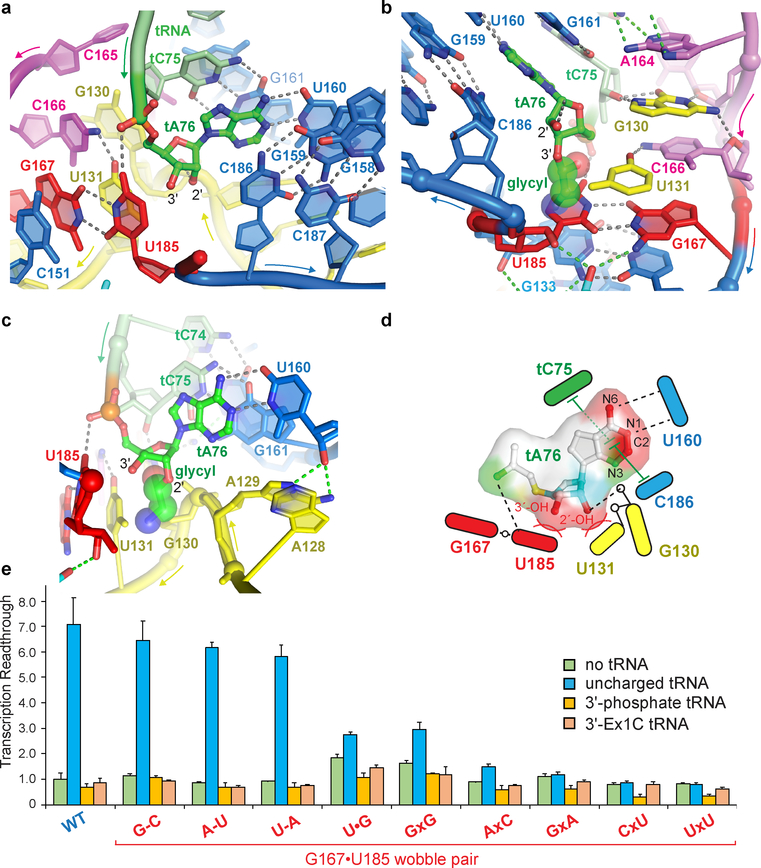
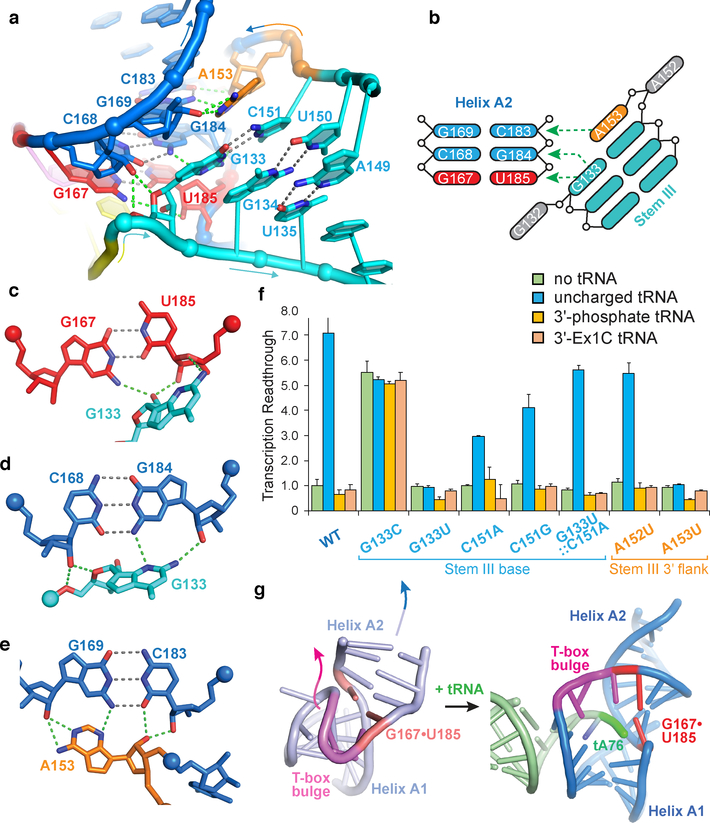
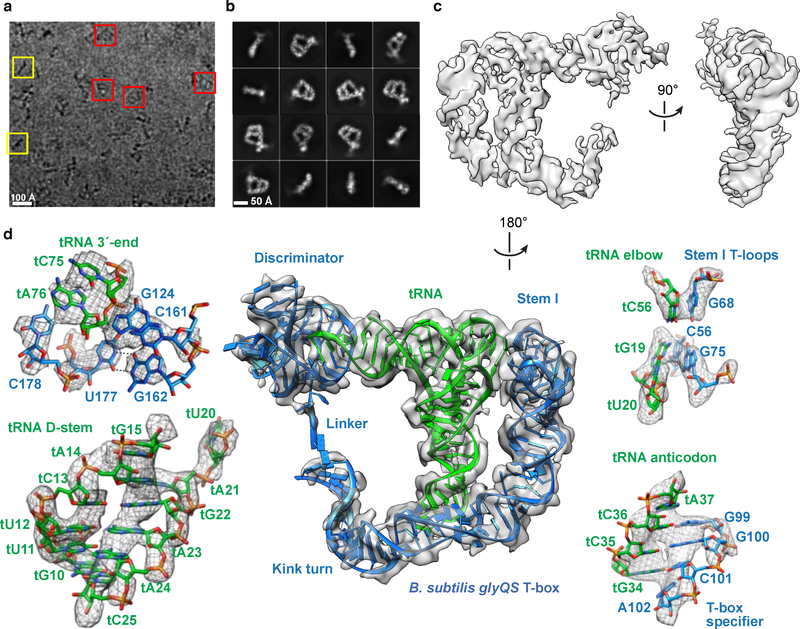
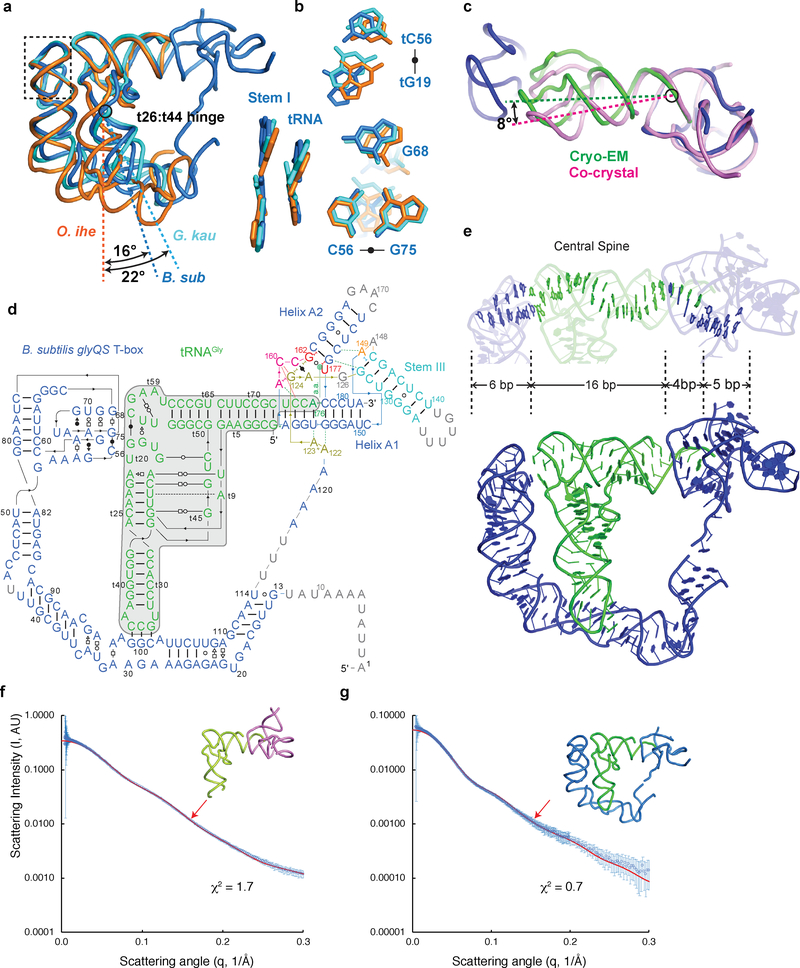
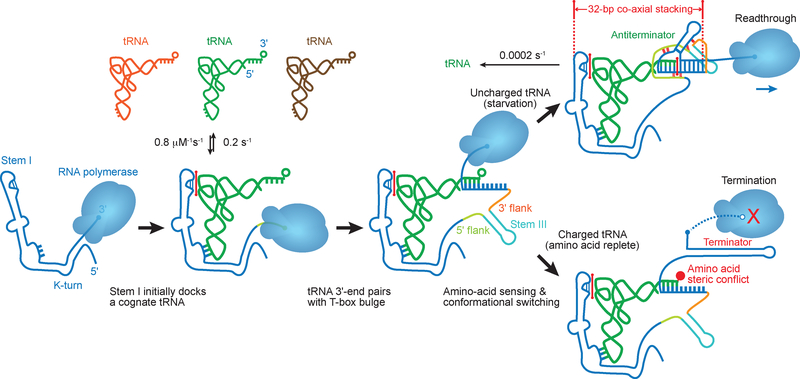
Amino acid availability in Gram-positive bacteria is monitored by T-box riboswitches. T-boxes directly bind tRNAs, assess their aminoacylation state, and regulate the transcription or translation of downstream genes to maintain nutritional homeostasis. Here, we report cocrystal and cryo-EM structures of Geobacillus kaustophilus and Bacillus subtilis T-box-tRNA complexes, detailing their multivalent, exquisitely selective interactions. The T-box forms a U-shaped molecular vise that clamps the tRNA, captures its 3' end using an elaborate 'discriminator' structure, and interrogates its aminoacylation state using a steric filter fashioned from a wobble base pair. In the absence of aminoacylation, T-boxes clutch tRNAs and form a continuously stacked central spine, permitting transcriptional readthrough or translation initiation. A modeled aminoacyl disrupts tRNA-T-box stacking, severing the central spine and blocking gene expression. Our data establish a universal mechanism of amino acid sensing on tRNAs and gene regulation by T-box riboswitches and exemplify how higher-order RNA-RNA interactions achieve multivalency and specificity.
Conflict of interest statement
Figures
Similar articles
-
An evolving tale of two interacting RNAs-themes and variations of the T-box riboswitch mechanism.IUBMB Life. 2019 Aug;71(8):1167-1180. doi: 10.1002/iub.2098. Epub 2019 Jun 17. IUBMB Life. 2019. PMID: 31206978 Free PMC article. Review.
-
Hierarchical mechanism of amino acid sensing by the T-box riboswitch.Nat Commun. 2018 May 14;9(1):1896. doi: 10.1038/s41467-018-04305-6. Nat Commun. 2018. PMID: 29760498 Free PMC article.
-
Capture and Release of tRNA by the T-Loop Receptor in the Function of the T-Box Riboswitch.Biochemistry. 2017 Jul 18;56(28):3549-3558. doi: 10.1021/acs.biochem.7b00284. Epub 2017 Jul 3. Biochemistry. 2017. PMID: 28621923 Free PMC article.
-
Direct evaluation of tRNA aminoacylation status by the T-box riboswitch using tRNA-mRNA stacking and steric readout.Mol Cell. 2014 Jul 3;55(1):148-55. doi: 10.1016/j.molcel.2014.05.017. Epub 2014 Jun 19. Mol Cell. 2014. PMID: 24954903 Free PMC article.
-
Unboxing the T-box riboswitches-A glimpse into multivalent and multimodal RNA-RNA interactions.Wiley Interdiscip Rev RNA. 2020 Nov;11(6):e1600. doi: 10.1002/wrna.1600. Epub 2020 Jul 6. Wiley Interdiscip Rev RNA. 2020. PMID: 32633085 Free PMC article. Review.
Cited by
-
Small-Molecule Antibiotics Inhibiting tRNA-Regulated Gene Expression Is a Viable Strategy for Targeting Gram-Positive Bacteria.Antimicrob Agents Chemother. 2020 Dec 16;65(1):e01247-20. doi: 10.1128/AAC.01247-20. Print 2020 Dec 16. Antimicrob Agents Chemother. 2020. PMID: 33077662 Free PMC article.
-
A riboswitch gives rise to multi-generational phenotypic heterogeneity in an auxotrophic bacterium.Nat Commun. 2020 Mar 5;11(1):1203. doi: 10.1038/s41467-020-15017-1. Nat Commun. 2020. PMID: 32139702 Free PMC article.
-
Recognition of the tRNA structure: Everything everywhere but not all at once.Cell Chem Biol. 2024 Jan 18;31(1):36-52. doi: 10.1016/j.chembiol.2023.12.008. Epub 2023 Dec 29. Cell Chem Biol. 2024. PMID: 38159570 Free PMC article. Review.
-
Cotranscriptional Assembly and Native Purification of Large RNA-RNA Complexes for Structural Analyses.Methods Mol Biol. 2023;2568:1-12. doi: 10.1007/978-1-0716-2687-0_1. Methods Mol Biol. 2023. PMID: 36227558 Free PMC article.
-
Interplay between Host tRNAs and HIV-1: A Structural Perspective.Viruses. 2021 Sep 13;13(9):1819. doi: 10.3390/v13091819. Viruses. 2021. PMID: 34578400 Free PMC article. Review.
References
-
- Antson AA et al. Structure of the trp RNA-binding attenuation protein, TRAP, bound to RNA. Nature 401, 235–42 (1999). - PubMed
Methods-only References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
