

Learning from methylomes: epigenomic correlates of Populus balsamifera traits based on deep learning models of natural DNA methylation
- PMID: 31742813
- PMCID: PMC7207000
- DOI: 10.1111/pbi.13299
Learning from methylomes: epigenomic correlates of Populus balsamifera traits based on deep learning models of natural DNA methylation
Abstract
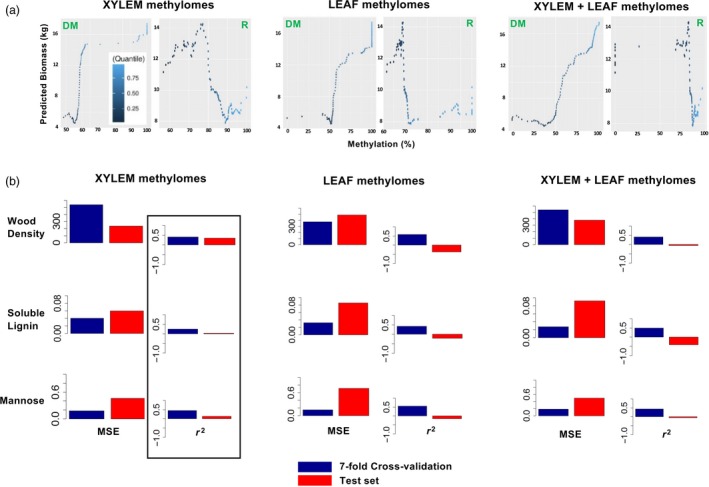
Epigenomes have remarkable potential for the estimation of plant traits. This study tested the hypothesis that natural variation in DNA methylation can be used to estimate industrially important traits in a genetically diverse population of Populus balsamifera L. (balsam poplar) trees grown at two common garden sites. Statistical learning experiments enabled by deep learning models revealed that plant traits in novel genotypes can be modelled transparently using small numbers of methylated DNA predictors. Using this approach, tissue type, a nonheritable attribute, from which DNA methylomes were derived was assigned, and provenance, a purely heritable trait and an element of population structure, was determined. Significant proportions of phenotypic variance in quantitative wood traits, including total biomass (57.5%), wood density (40.9%), soluble lignin (25.3%) and cell wall carbohydrate (mannose: 44.8%) contents, were also explained from natural variation in DNA methylation. Modelling plant traits using DNA methylation can capture tissue-specific epigenetic mechanisms underlying plant phenotypes in natural environments. DNA methylation-based models offer new insight into natural epigenetic influence on plants and can be used as a strategy to validate the identity, provenance or quality of agroforestry products.
Keywords: authentication; deep learning; epigenomics; poplar.
© 2019 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflicts of interest related to this work.
Figures







References
-
- Alipanahi, B. , Delong, A. , Weirauch, M.T. and Frey, B.J. (2015) Predicting the sequence specificities of DNA‐ and RNA‐binding proteins by deep learning. Nat Biotechnol. 33, 831–838. - PubMed
-
- Bengio, Y. (1998) Practical recommendations for gradient-based training of deep architectures In Neural Networks: Tricks of the Trade (Montavon G., Orr G.B. and Müller K.R. eds), pp. 437–478. Berlin: Springer.
Publication types
MeSH terms
Associated data
- Actions
LinkOut - more resources
Full Text Sources
