

Tricalbin-Mediated Contact Sites Control ER Curvature to Maintain Plasma Membrane Integrity
- PMID: 31743662
- PMCID: PMC6863395
- DOI: 10.1016/j.devcel.2019.10.018
Tricalbin-Mediated Contact Sites Control ER Curvature to Maintain Plasma Membrane Integrity
Abstract
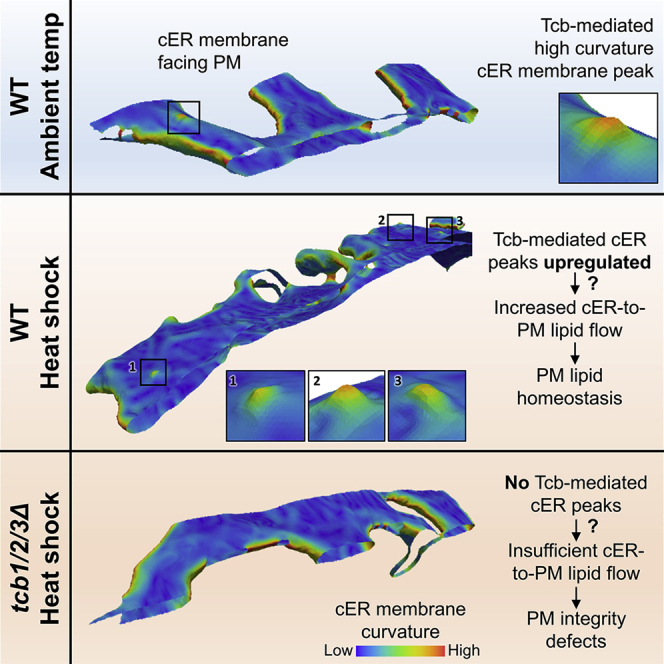
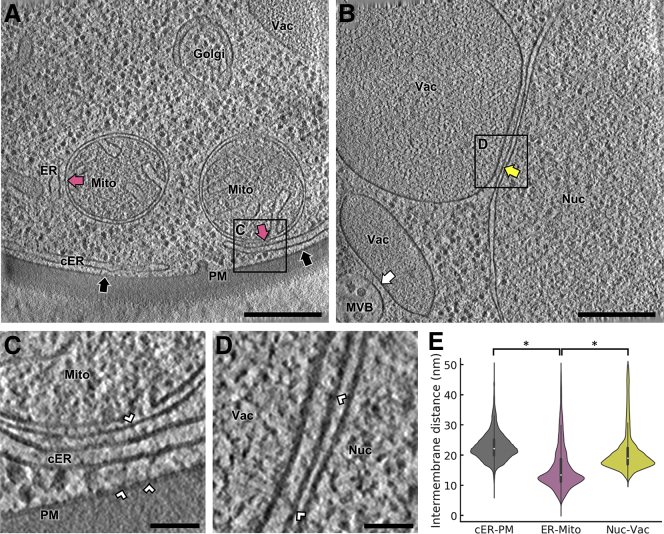
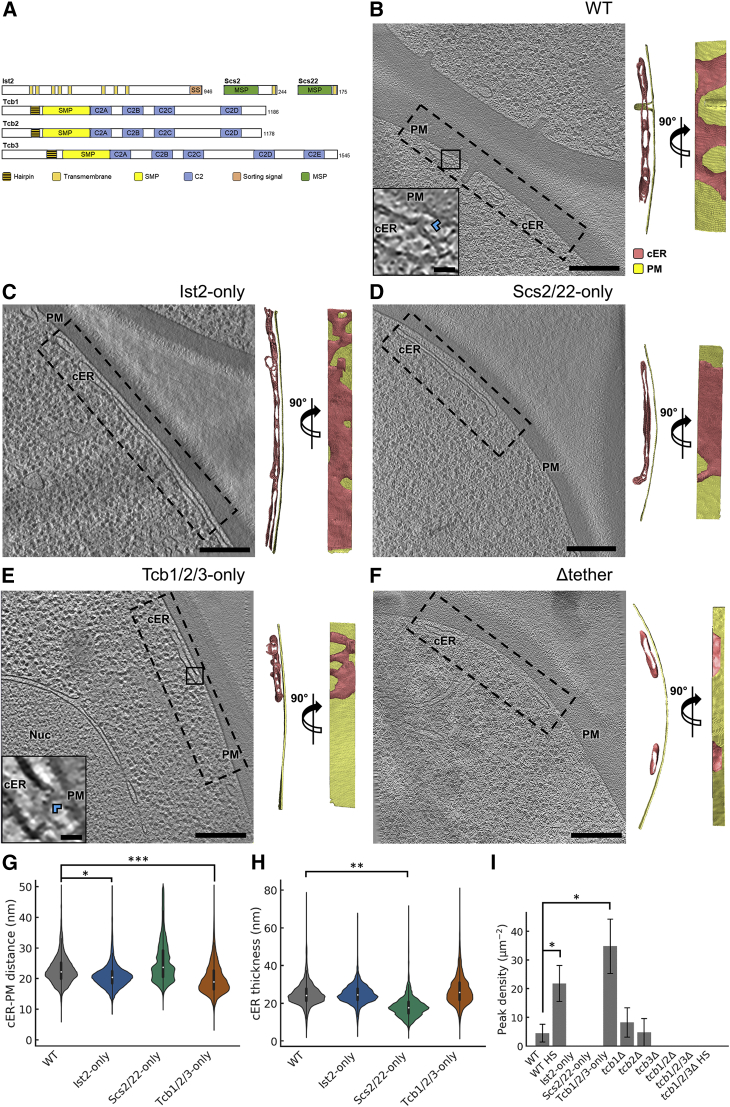
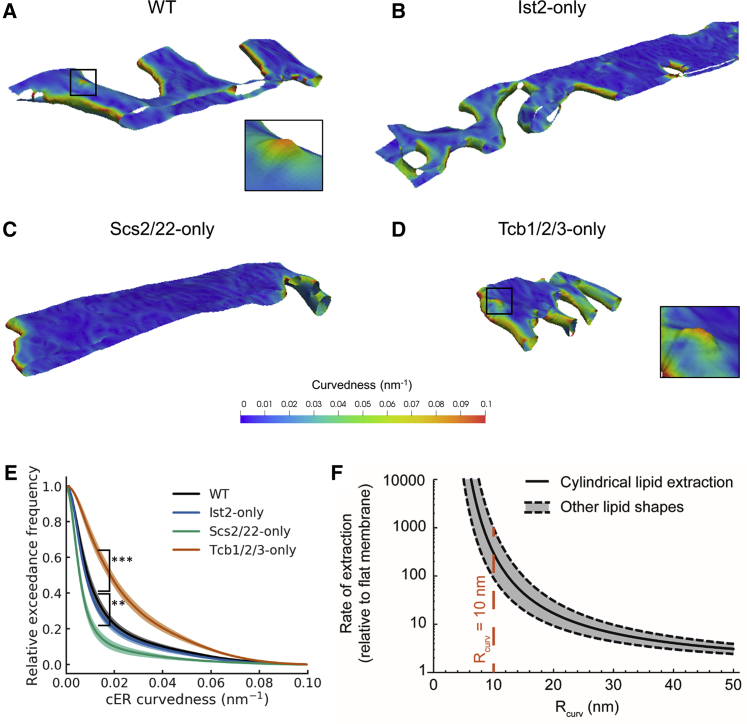
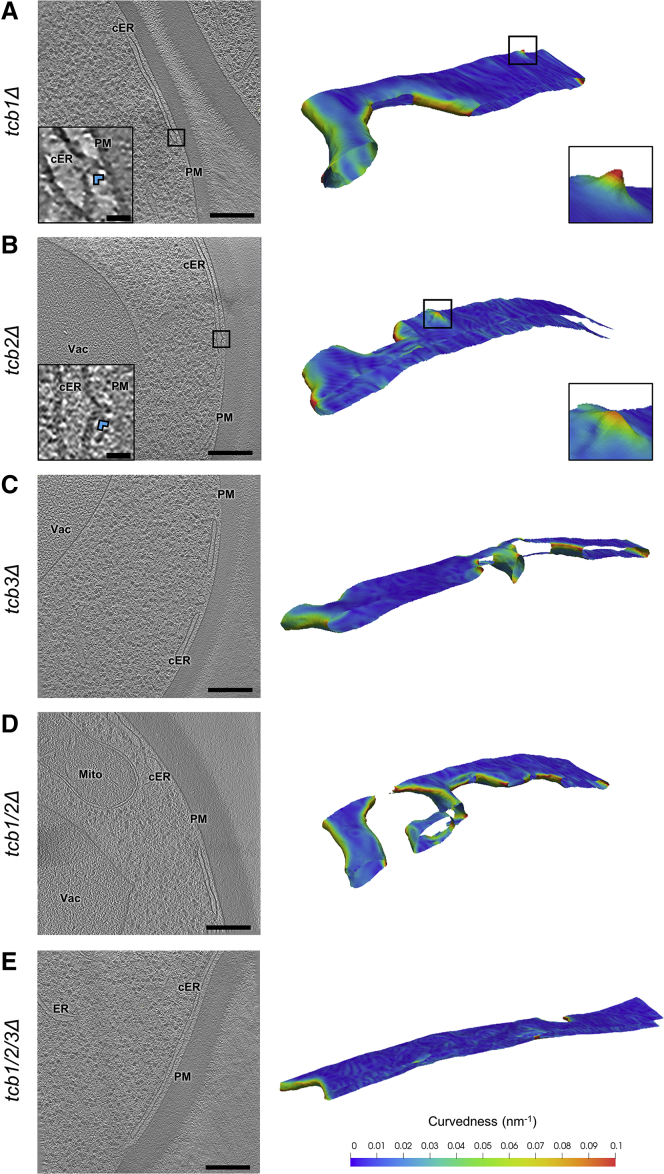
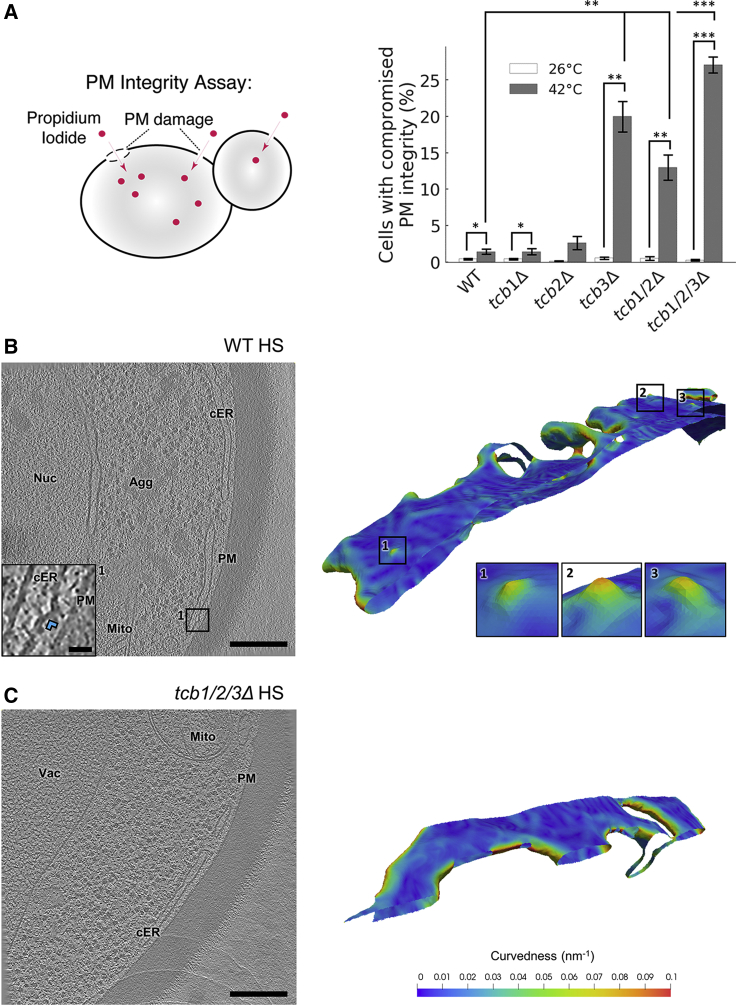
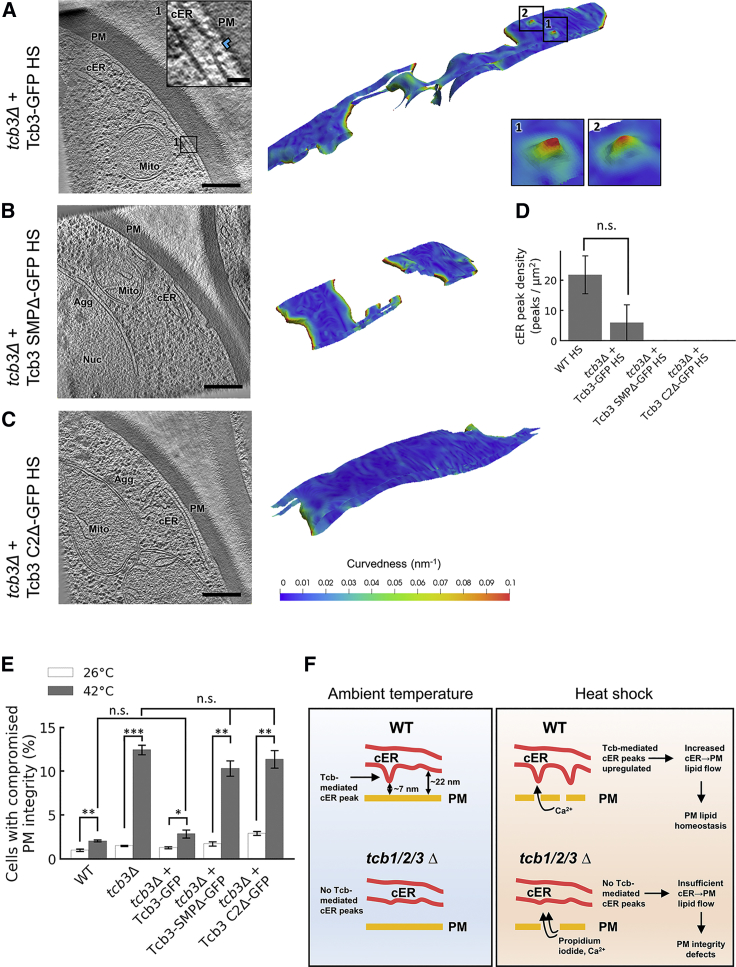
Membrane contact sites (MCS) between the endoplasmic reticulum (ER) and the plasma membrane (PM) play fundamental roles in all eukaryotic cells. ER-PM MCS are particularly abundant in Saccharomyces cerevisiae, where approximately half of the PM surface is covered by cortical ER (cER). Several proteins, including Ist2, Scs2/22, and Tcb1/2/3 are implicated in cER formation, but the specific roles of these molecules are poorly understood. Here, we use cryo-electron tomography to show that ER-PM tethers are key determinants of cER morphology. Notably, Tcb proteins (tricalbins) form peaks of extreme curvature on the cER membrane facing the PM. Combined modeling and functional assays suggest that Tcb-mediated cER peaks facilitate the transport of lipids between the cER and the PM, which is necessary to maintain PM integrity under heat stress. ER peaks were also present at other MCS, implying that membrane curvature enforcement may be a widespread mechanism to regulate MCS function.
Keywords: cryo-ET; cryo-FIB; cryo-electron tomography; cryo-focused ion beam milling; endoplasmic reticulum; membrane contact site; membrane curvature; plasma membrane; tricalbins.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Ahrens J., Geveci B., Law C. Visualization Handbook. Elsevier; 2005. ParaView: an end-user tool for large data visualization.
-
- Andrews N.W., Corrotte M. Plasma membrane repair. Curr. Biol. 2018;28:R392–R397. - PubMed
-
- Antonny B. Mechanisms of membrane curvature sensing. Annu. Rev. Biochem. 2011;80:101–123. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
