

Atypical organizations and epistatic interactions of CRISPRs and cas clusters in genomes and their mobile genetic elements
- PMID: 31745554
- PMCID: PMC7145637
- DOI: 10.1093/nar/gkz1091
Atypical organizations and epistatic interactions of CRISPRs and cas clusters in genomes and their mobile genetic elements
Abstract
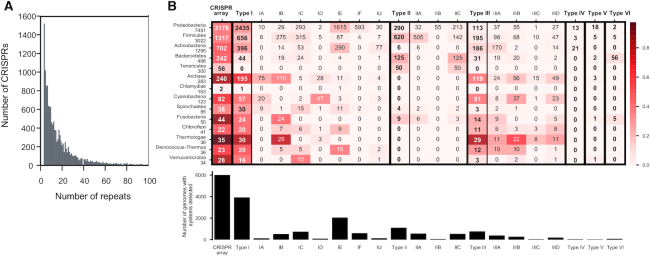
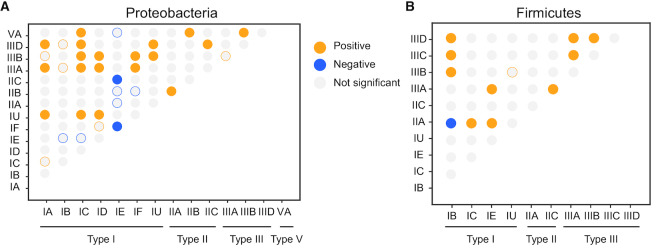
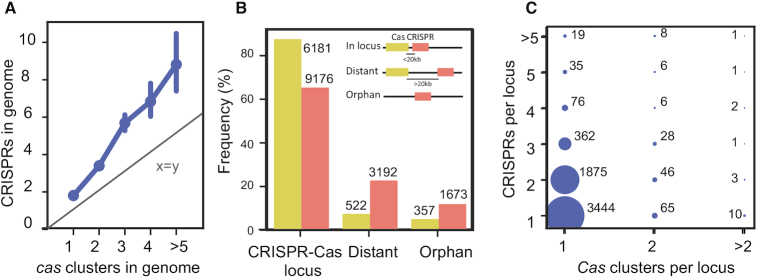
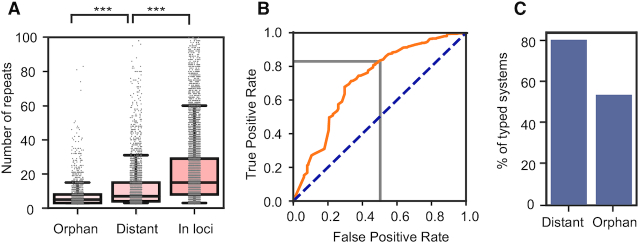
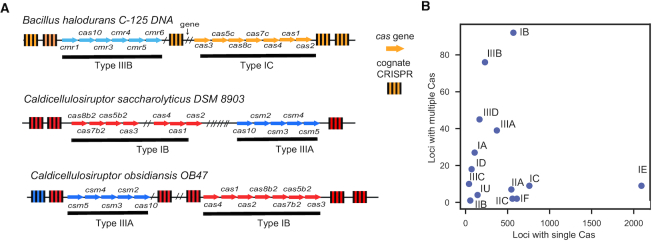
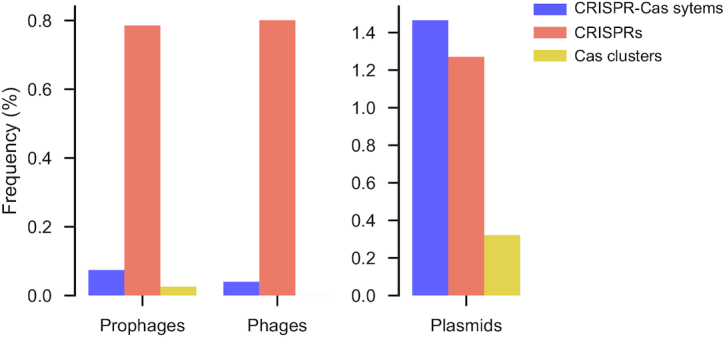
Prokaryotes use CRISPR-Cas systems for adaptive immunity, but the reasons for the frequent existence of multiple CRISPRs and cas clusters remain poorly understood. Here, we analysed the joint distribution of CRISPR and cas genes in a large set of fully sequenced bacterial genomes and their mobile genetic elements. Our analysis suggests few negative and many positive epistatic interactions between Cas subtypes. The latter often result in complex genetic organizations, where a locus has a single adaptation module and diverse interference mechanisms that might provide more effective immunity. We typed CRISPRs that could not be unambiguously associated with a cas cluster and found that such complex loci tend to have unique type I repeats in multiple CRISPRs. Many chromosomal CRISPRs lack a neighboring Cas system and they often have repeats compatible with the Cas systems encoded in trans. Phages and 25 000 prophages were almost devoid of CRISPR-Cas systems, whereas 3% of plasmids had CRISPR-Cas systems or isolated CRISPRs. The latter were often compatible with the chromosomal cas clusters, suggesting that plasmids can co-opt the latter. These results highlight the importance of interactions between CRISPRs and cas present in multiple copies and in distinct genomic locations in the function and evolution of bacterial immunity.
© The Author(s) 2019. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Barrangou R., Fremaux C., Deveau H., Richards M., Boyaval P., Moineau S., Romero D.A., Horvath P.. CRISPR provides acquired resistance against viruses in prokaryotes. Science. 2007; 315:1709–1712. - PubMed
-
- Garneau J.E., Dupuis M.-È., Villion M., Romero D.A., Barrangou R., Boyaval P., Fremaux C., Horvath P., Magadán A.H., Moineau S.. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature. 2010; 468:67–71. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
