

Alterations in sleep, sleep spindle, and EEG power in mGluR5 knockout mice
- PMID: 31747354
- PMCID: PMC6985862
- DOI: 10.1152/jn.00532.2019
Alterations in sleep, sleep spindle, and EEG power in mGluR5 knockout mice
Abstract
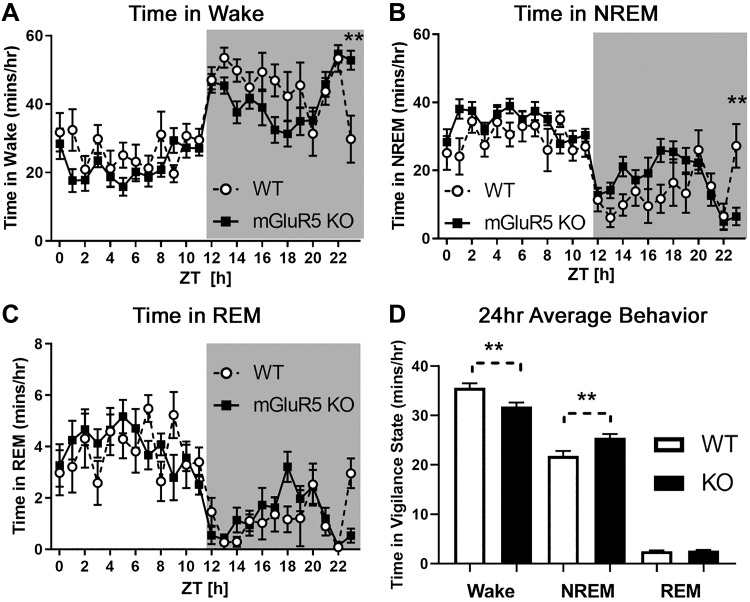
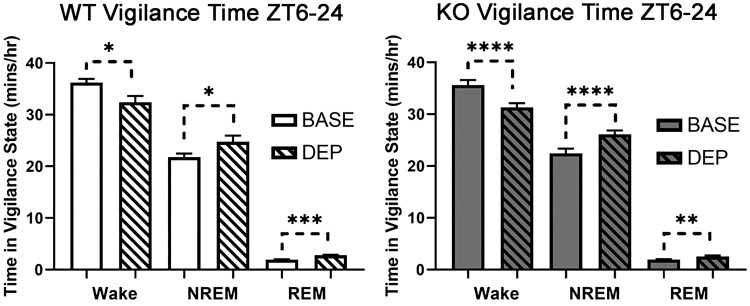
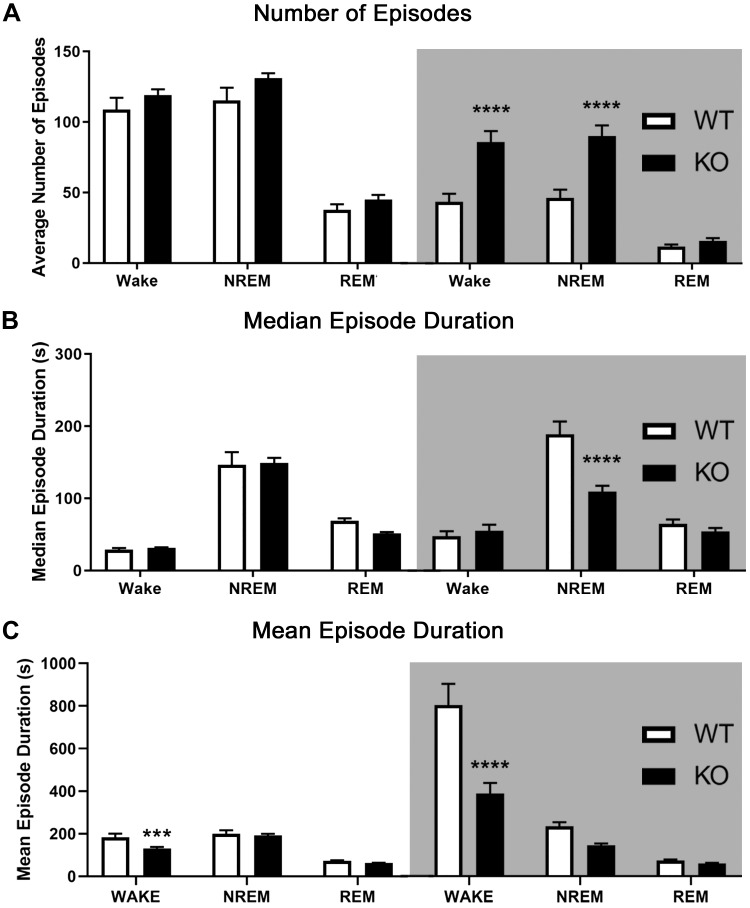
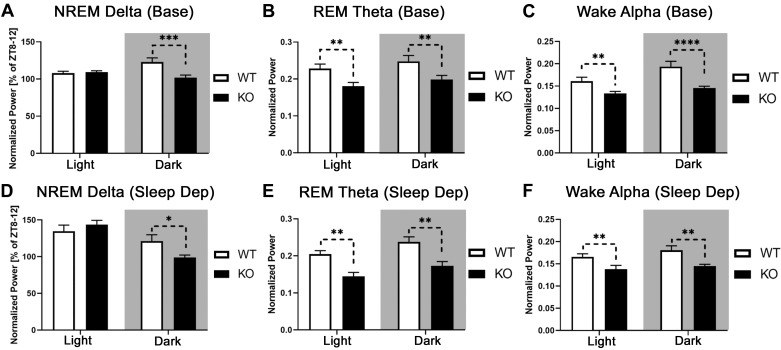
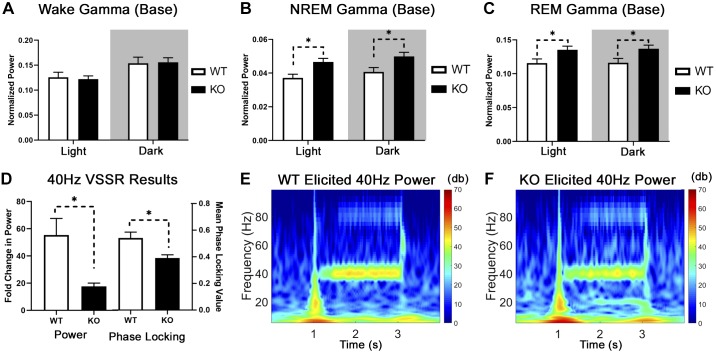
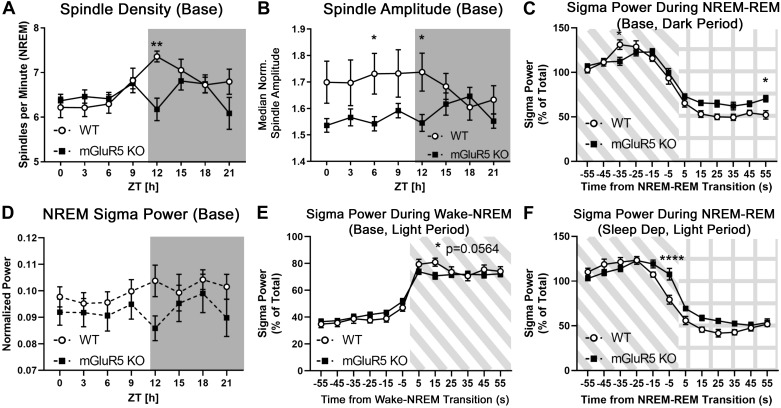
The type 5 metabotropic glutamate receptor (mGluR5) represents a novel therapeutic target for schizophrenia and other disorders. Schizophrenia is associated with progressive abnormalities in cortical oscillatory processes including reduced spindles (8-15 Hz) during sleep and increased delta (0.5-4 Hz)- and gamma-band activity (30-80 Hz) during wakefulness. mGluR5 knockout (KO) mice demonstrate many schizophrenia-like behaviors, including abnormal sleep. To examine the effects of mGluR5 on the maintenance of the neocortical circuitry responsible for such neural oscillations, we analyzed sleep/wake electroencephalographic (EEG) activity of mGluR5 KO mice at baseline, after 6 h of sleep deprivation, and during a visual method of cortical entrainment (visual steady state response). We hypothesized mGluR5-KO mice would exhibit translationally relevant abnormalities in sleep and neural oscillations that mimic schizophrenia. Power spectral and spindle density analyses were performed across 24-h EEG recordings in mGluR5-KO mice and wild-type (WT) controls. Novel findings in mGluR5 KO mice include deficits in sleep spindle density, wake alpha power, and 40-Hz visual task-evoked gamma power and phase locking. Sigma power (10-15 Hz), an approximation of spindle activity, was also reduced during non-rapid eye movement sleep transitions. Our observations on abnormal sleep/wake are generally in agreement with previous reports, although we did not replicate changes in rapid eye movement sleep. The timing of these phenotypes may suggest an impaired circadian process in mGluR5 KO mice. In conclusion, EEG phenotypes in mGluR5 KO mice resemble deficits observed in patients with schizophrenia. These findings implicate mGluR5-mediated pathways in several translationally relevant phenotypes associated with schizophrenia, and suggest that agents targeting this receptor may have harmful consequences on sleep health and daily patterns of EEG power.NEW & NOTEWORTHY Metabotropic glutamate receptor type 5 (mGluR5) knockout (KO) mice show several translationally relevant abnormalities in neural oscillatory activity associated with schizophrenia. These include deficits in sleep spindle density, sigma and alpha power, and 40-Hz task-evoked gamma power. The timing of these phenotypes suggests an impaired circadian process in these mice. Previously reported rapid eye movement sleep deficits in this model were not observed. These findings suggest mGluR5-enhancing drugs may improve sleep stability and sleep spindle density, which could impact memory and cognition.
Keywords: N-methyl-d-aspartate receptors; metabotropic glutamate receptor 5; polysomnography; sigma; visual evoked potentials.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures
References
-
- Ahnaou A, Langlois X, Steckler T, Bartolome-Nebreda JM, Drinkenburg WH. Negative versus positive allosteric modulation of metabotropic glutamate receptors (mGluR5): indices for potential pro-cognitive drug properties based on EEG network oscillations and sleep-wake organization in rats. Psychopharmacology (Berl) 232: 1107–1122, 2015a. doi:10.1007/s00213-014-3746-4. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials