

Occurrence and Functions of m6A and Other Covalent Modifications in Plant mRNA
- PMID: 31748418
- PMCID: PMC6945878
- DOI: 10.1104/pp.19.01156
Occurrence and Functions of m6A and Other Covalent Modifications in Plant mRNA
Abstract
Posttranscriptional control of gene expression is indispensable for the execution of developmental programs and environmental adaptation. Among the many cellular mechanisms that regulate mRNA fate, covalent nucleotide modification has emerged as a major way of controlling the processing, localization, stability, and translatability of mRNAs. This powerful mechanism is conserved across eukaryotes and controls the cellular events that lead to development and growth. As in other eukaryotes, N 6-methylation of adenosine is the most abundant and best studied mRNA modification in flowering plants. It is essential for embryonic and postembryonic plant development and it affects growth rate and stress responses, including susceptibility to plant RNA viruses. Although the mRNA modification field is young, the intense interest triggered by its involvement in stem cell differentiation and cancer has led to rapid advances in understanding how mRNA modifications control gene expression in mammalian systems. An equivalent effort from plant molecular biologists has been lagging behind, but recent work in Arabidopsis (Arabidopsis thaliana) and other plant species is starting to give insights into how this essential layer of posttranscriptional regulation works in plants, and both similarities and differences with other eukaryotes are emerging. In this Update, we summarize, connect, and evaluate the experimental work that supports our current knowledge of the biochemistry, molecular mechanisms, and biological functions of mRNA modifications in plants. We devote particular attention to N 6-methylation of adenosine and attempt to place the knowledge gained from plant studies within the context of a more general framework derived from studies in other eukaryotes.
© 2020 American Society of Plant Biologists. All Rights Reserved.
Figures
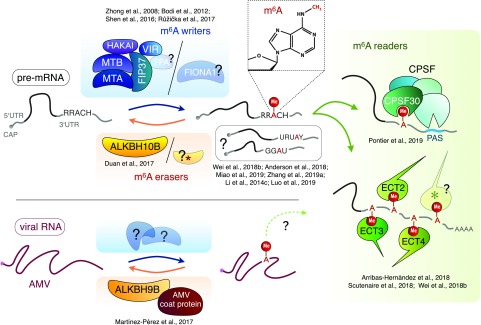
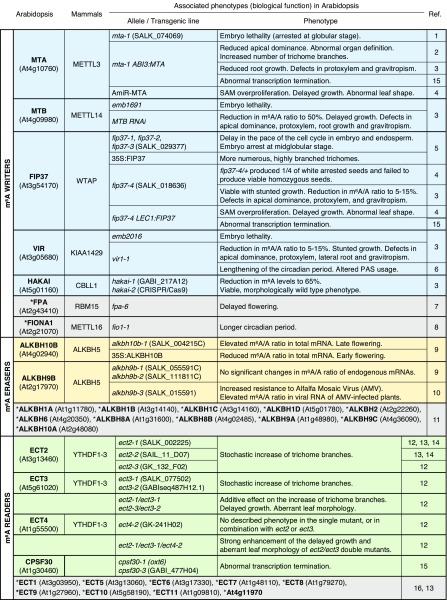
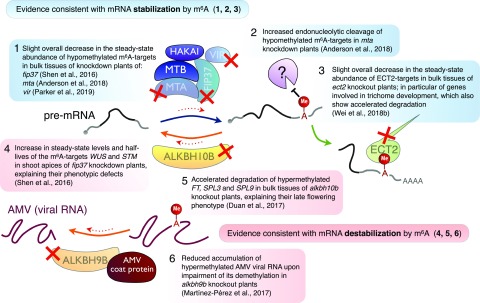
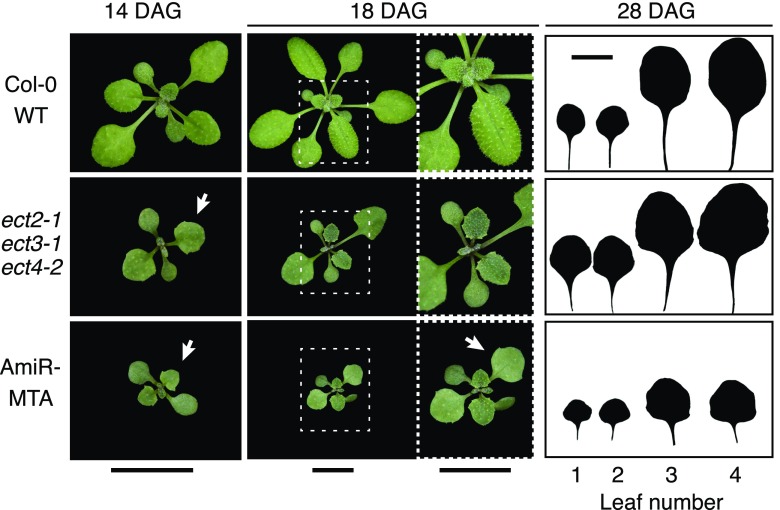





References
-
- Anderson SJ, Kramer MC, Gosai SJ, Yu X, Vandivier LE, Nelson ADL, Anderson ZD, Beilstein MA, Fray RG, Lyons E, et al. (2018) N6-Methyladenosine inhibits local ribonucleolytic cleavage to stabilize mRNAs in Arabidopsis. Cell Rep 25: 1146–1157 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
