

PGRMC2 is an intracellular haem chaperone critical for adipocyte function
- PMID: 31748741
- PMCID: PMC6895438
- DOI: 10.1038/s41586-019-1774-2
PGRMC2 is an intracellular haem chaperone critical for adipocyte function
Abstract
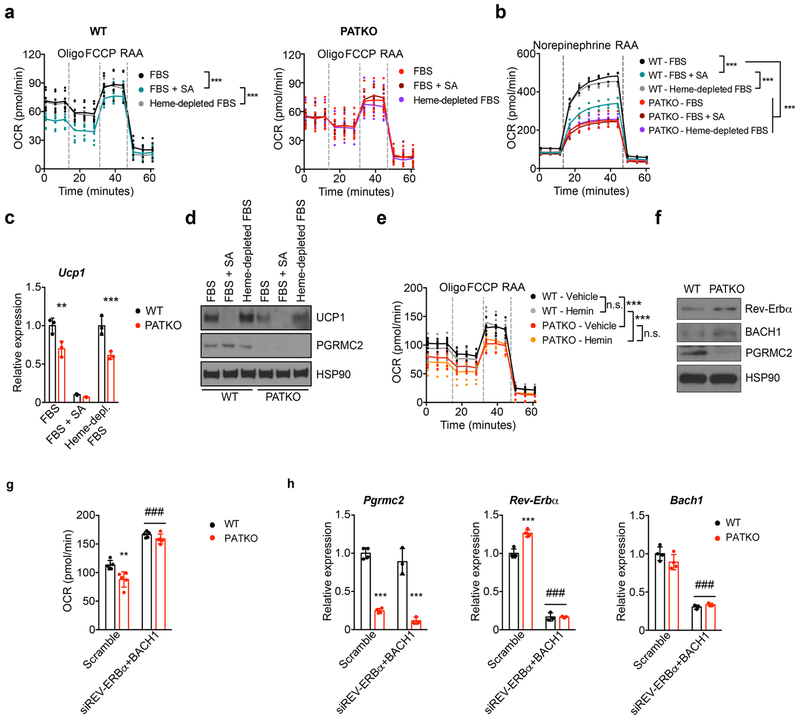
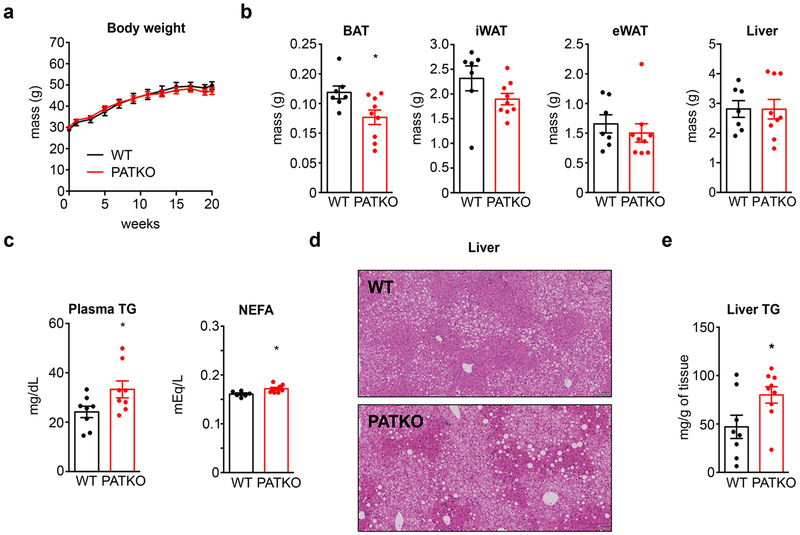
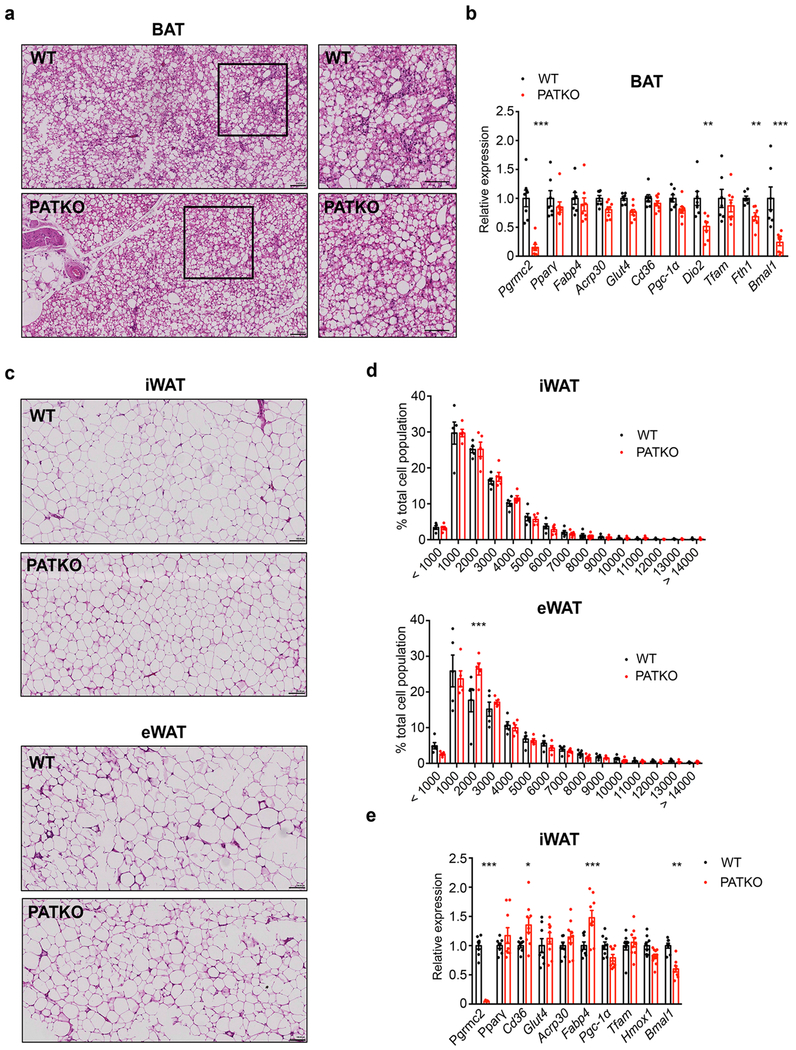
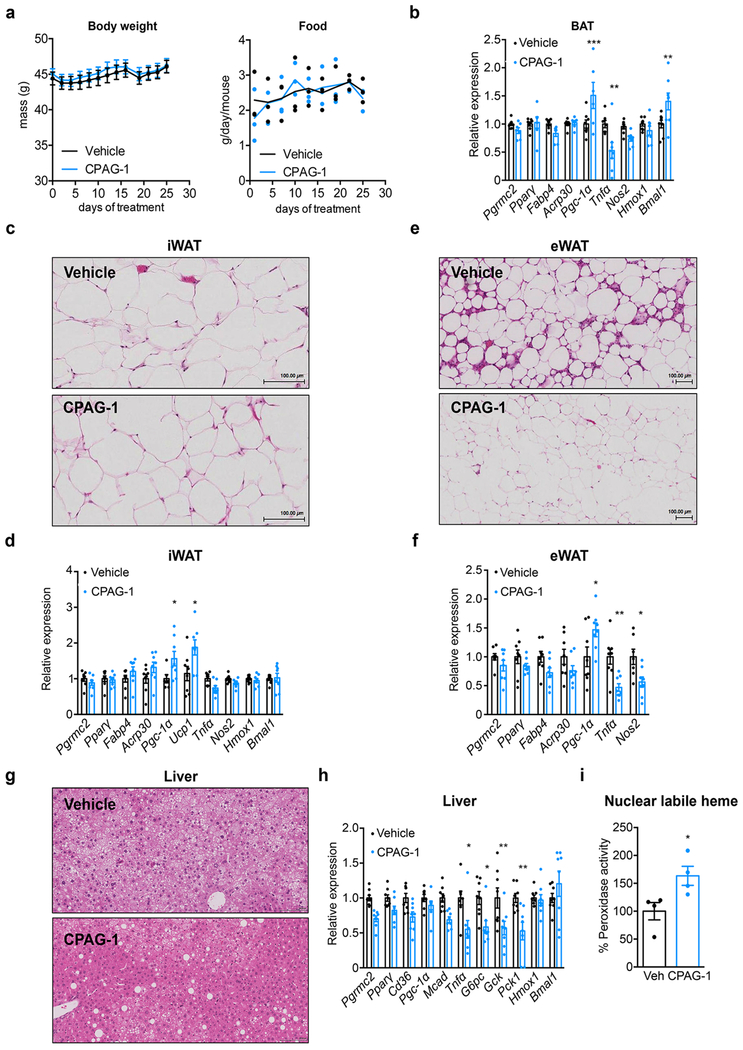
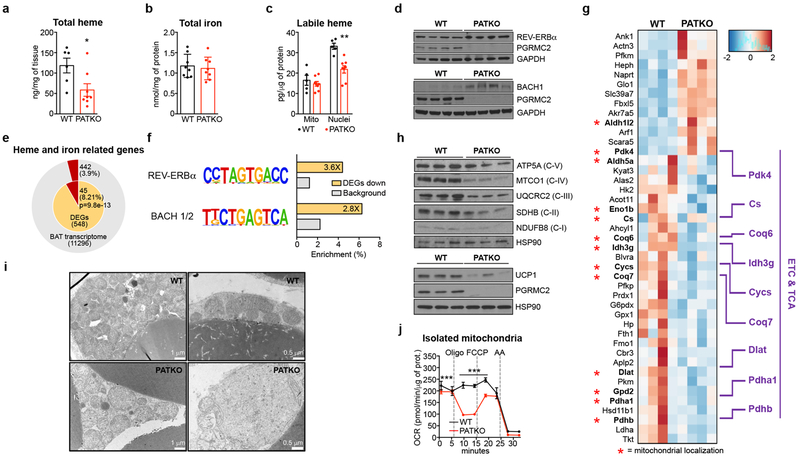
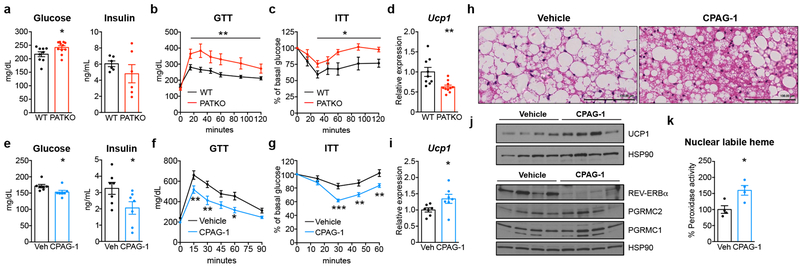
Haem is an essential prosthetic group of numerous proteins and a central signalling molecule in many physiologic processes1,2. The chemical reactivity of haem means that a network of intracellular chaperone proteins is required to avert the cytotoxic effects of free haem, but the constituents of such trafficking pathways are unknown3,4. Haem synthesis is completed in mitochondria, with ferrochelatase adding iron to protoporphyrin IX. How this vital but highly reactive metabolite is delivered from mitochondria to haemoproteins throughout the cell remains poorly defined3,4. Here we show that progesterone receptor membrane component 2 (PGRMC2) is required for delivery of labile, or signalling haem, to the nucleus. Deletion of PGMRC2 in brown fat, which has a high demand for haem, reduced labile haem in the nucleus and increased stability of the haem-responsive transcriptional repressors Rev-Erbα and BACH1. Ensuing alterations in gene expression caused severe mitochondrial defects that rendered adipose-specific PGRMC2-null mice unable to activate adaptive thermogenesis and prone to greater metabolic deterioration when fed a high-fat diet. By contrast, obese-diabetic mice treated with a small-molecule PGRMC2 activator showed substantial improvement of diabetic features. These studies uncover a role for PGRMC2 in intracellular haem transport, reveal the influence of adipose tissue haem dynamics on physiology and suggest that modulation of PGRMC2 may revert obesity-linked defects in adipocytes.
Conflict of interest statement
Figures
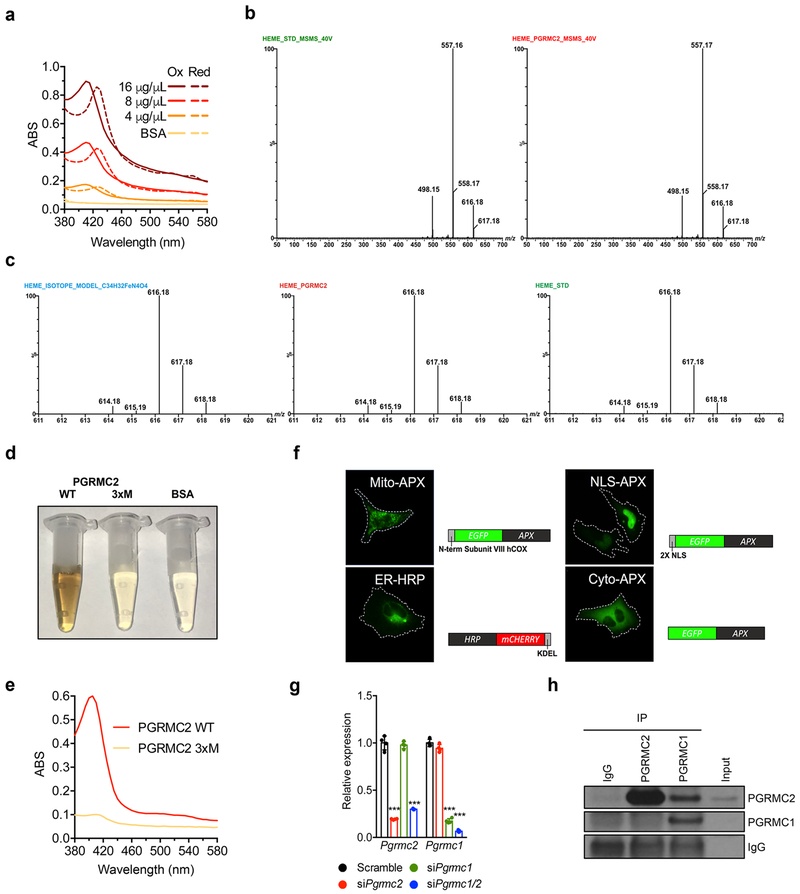
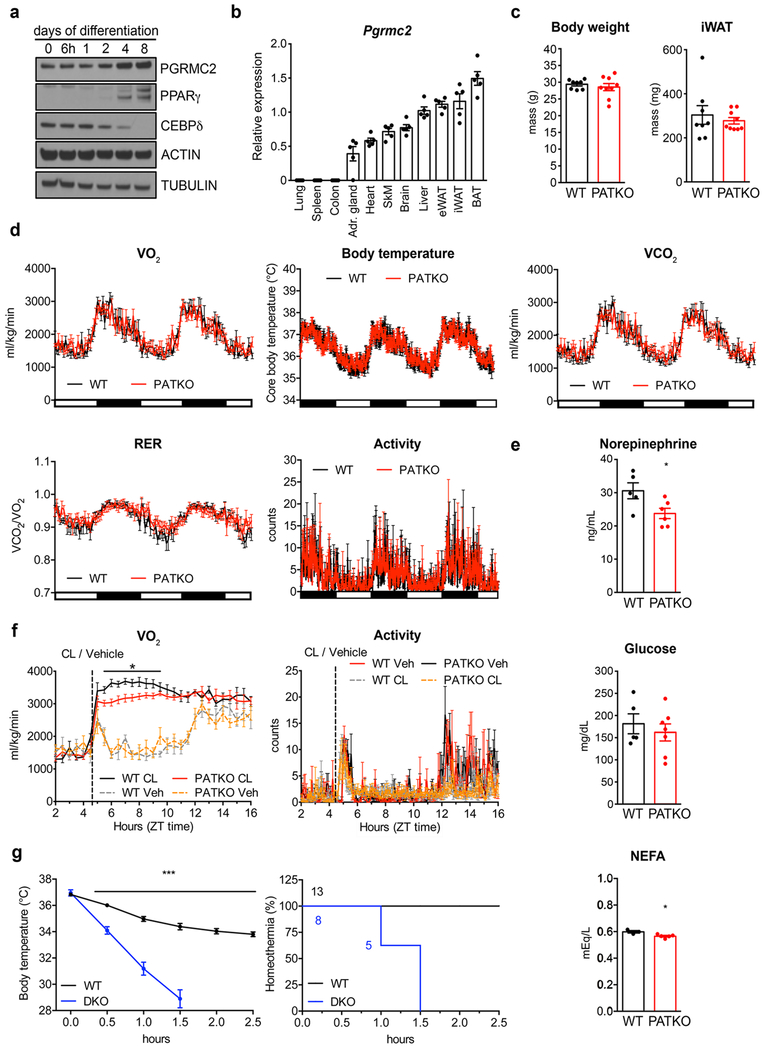
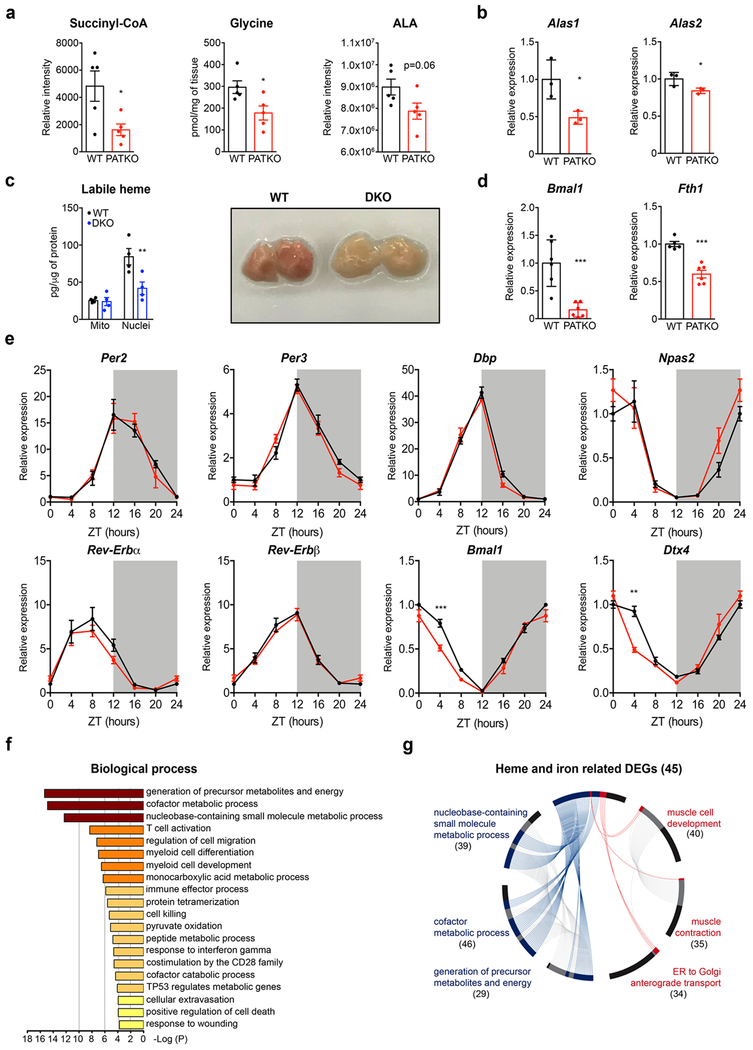
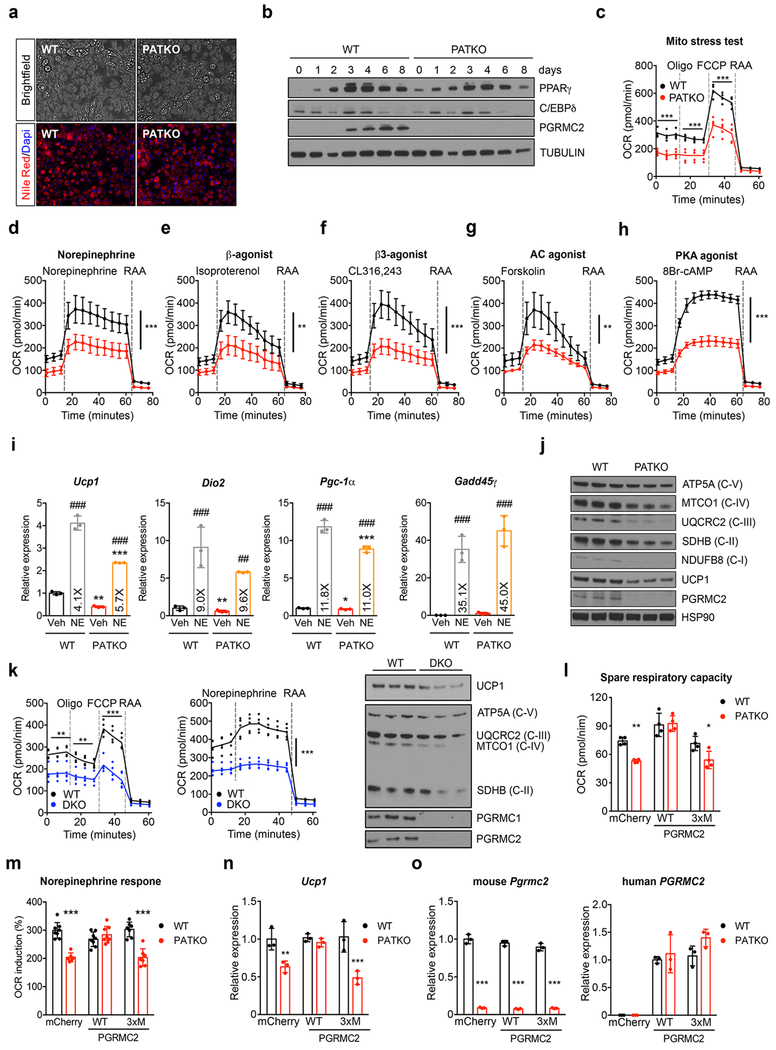
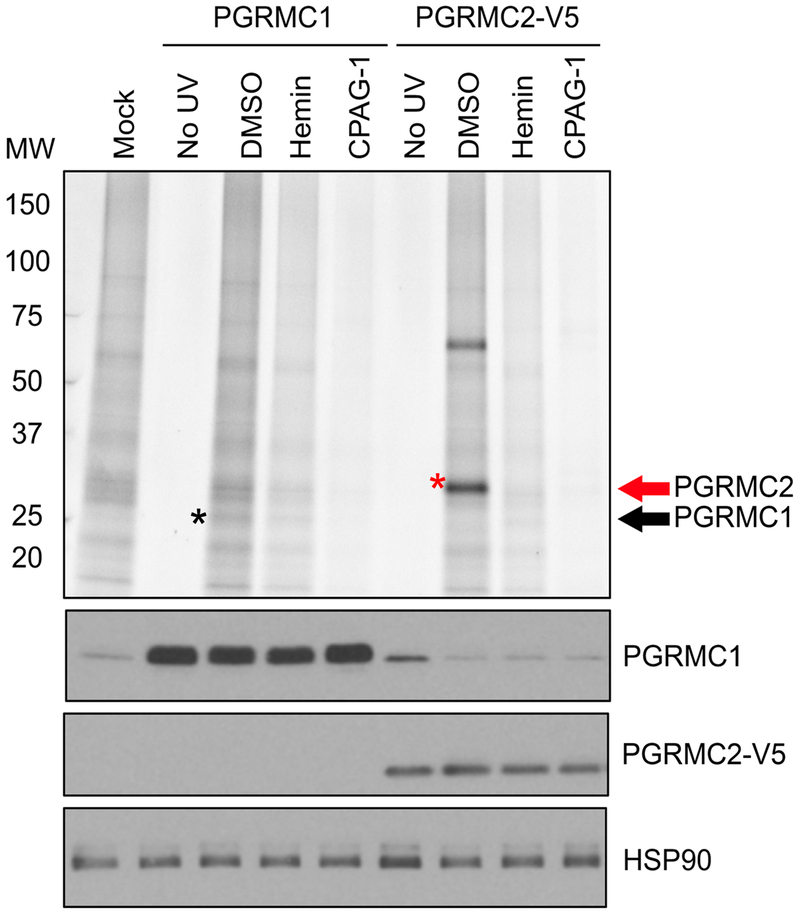
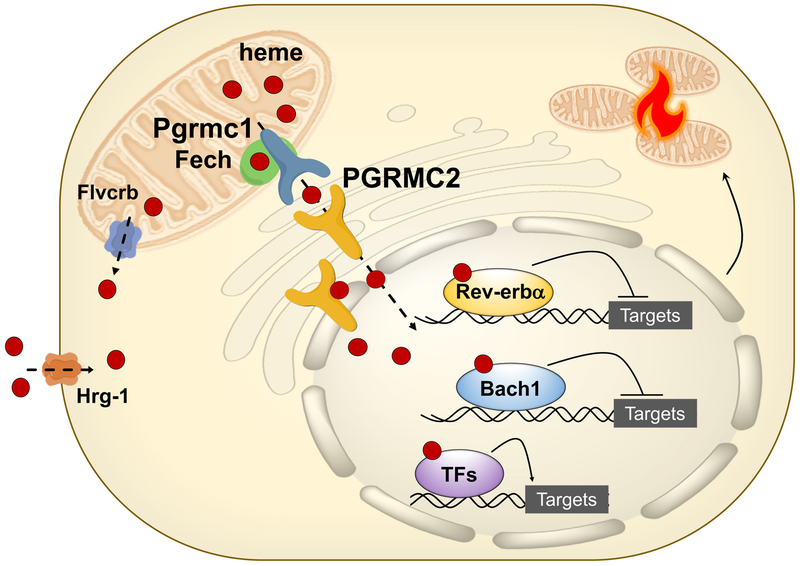
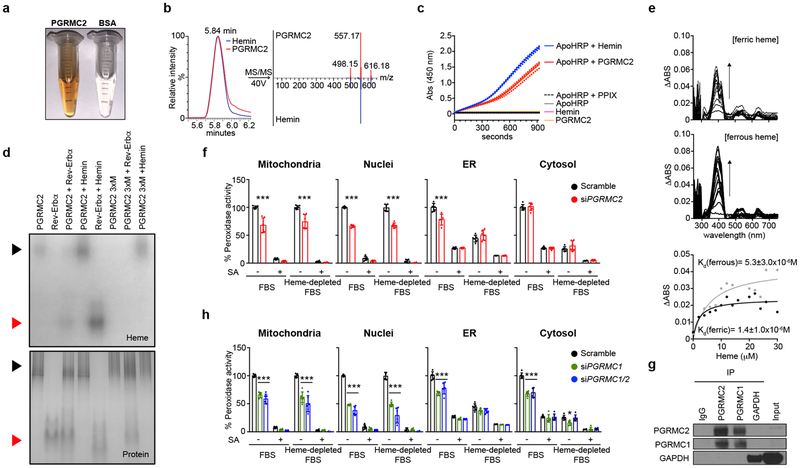
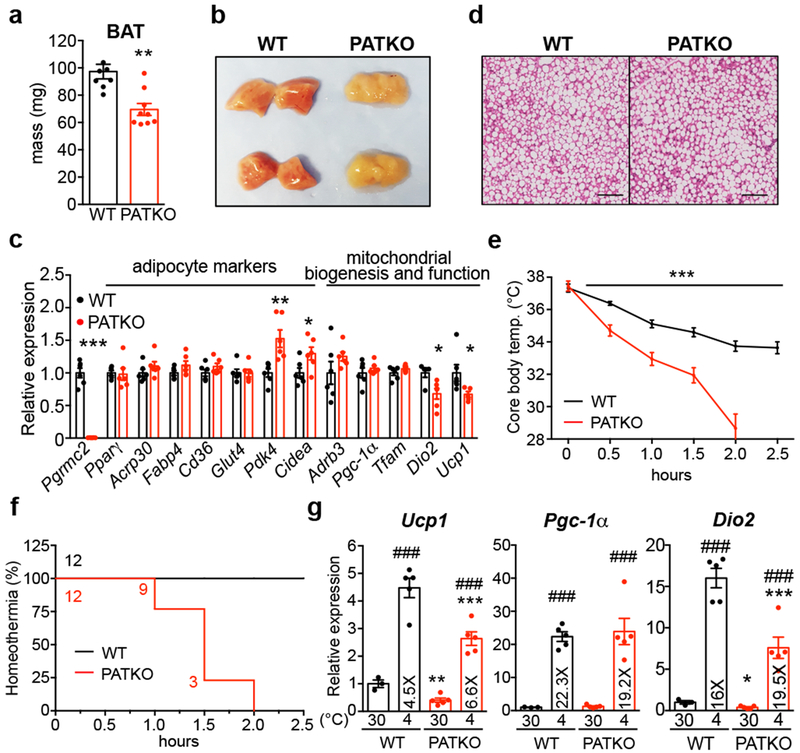
References
-
- Mense SM & Zhang L Heme: a versatile signaling molecule controlling the activities of diverse regulators ranging from transcription factors to MAP kinases. Cell Res 16, 681–692 (2006). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
