

Nickel Toxicity Targets Cell Wall-Related Processes and PIN2-Mediated Auxin Transport to Inhibit Root Elongation and Gravitropic Responses in Arabidopsis
- PMID: 31750920
- PMCID: PMC7065594
- DOI: 10.1093/pcp/pcz217
Nickel Toxicity Targets Cell Wall-Related Processes and PIN2-Mediated Auxin Transport to Inhibit Root Elongation and Gravitropic Responses in Arabidopsis
Abstract
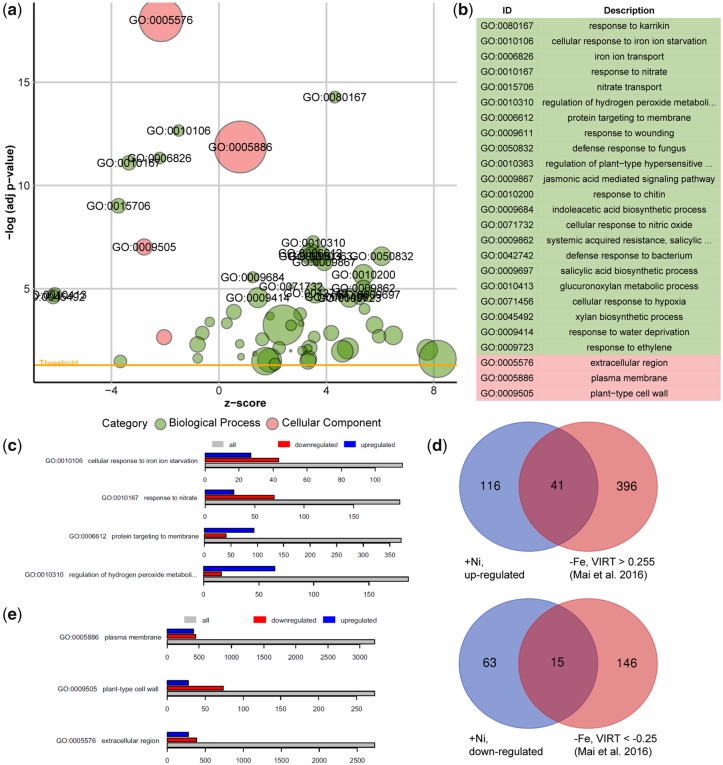
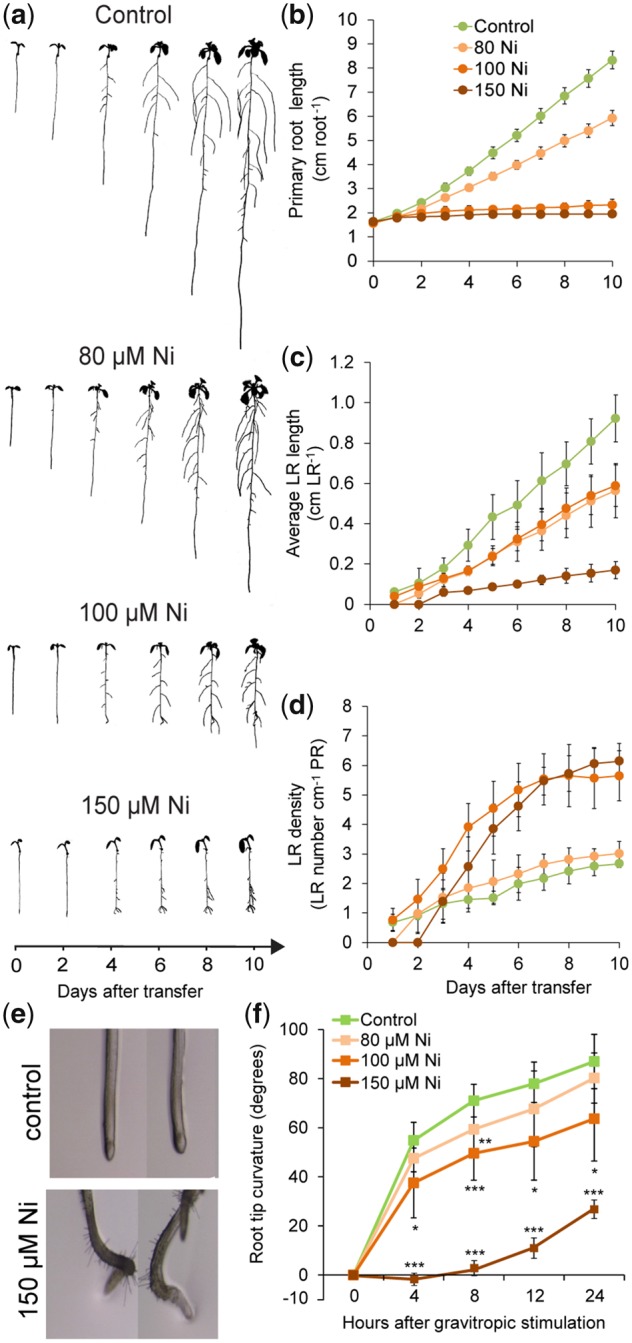
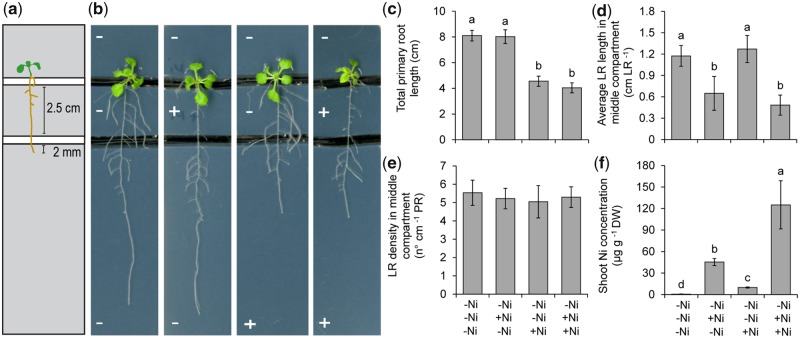
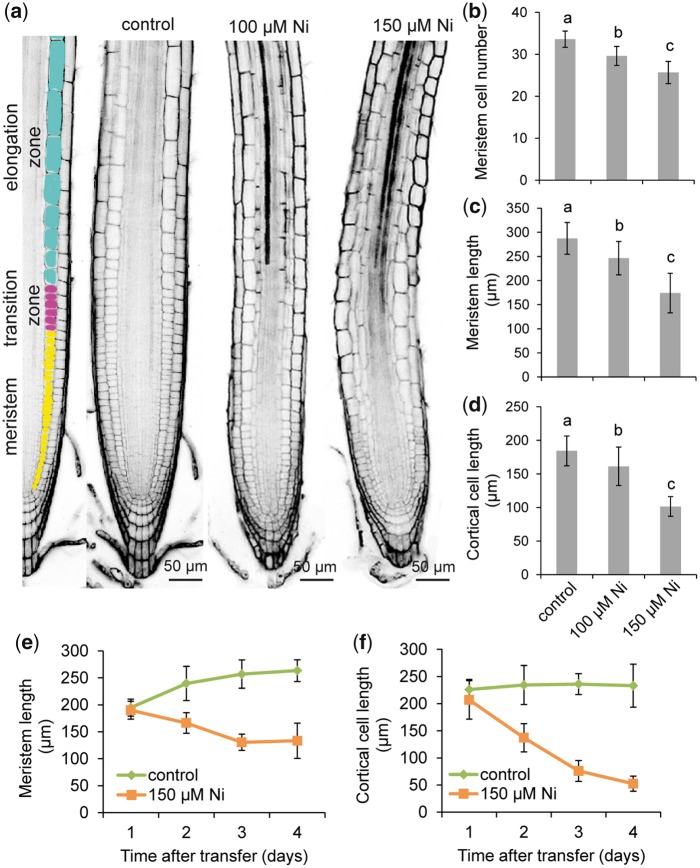
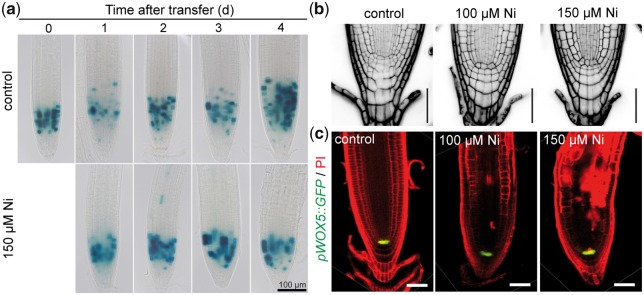
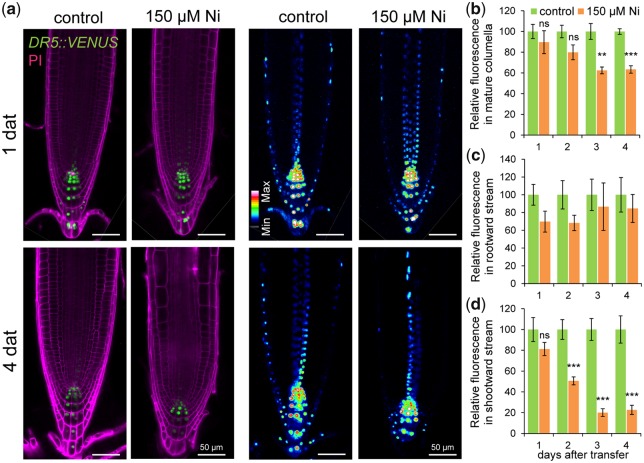
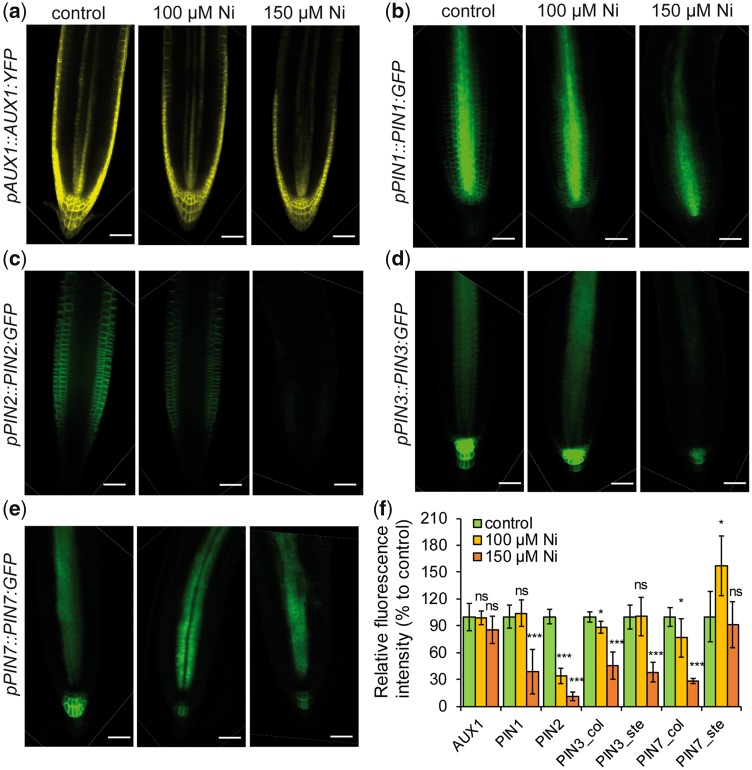
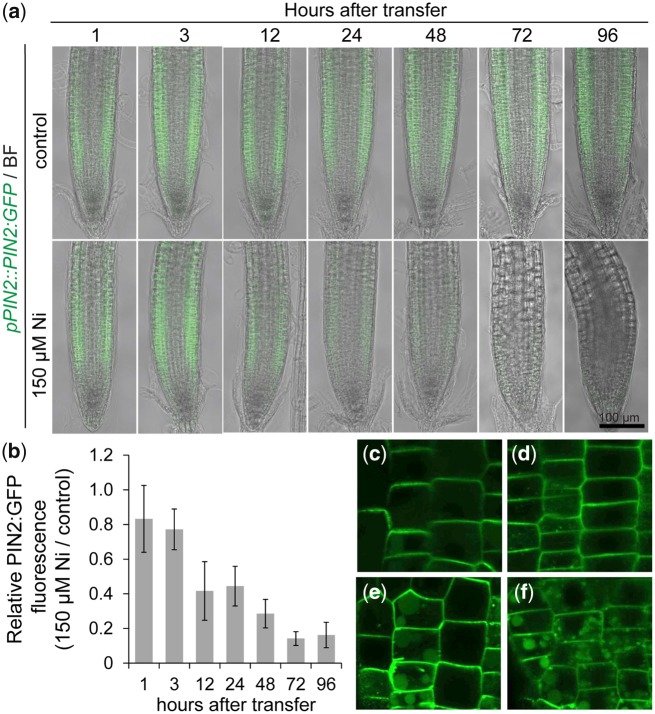
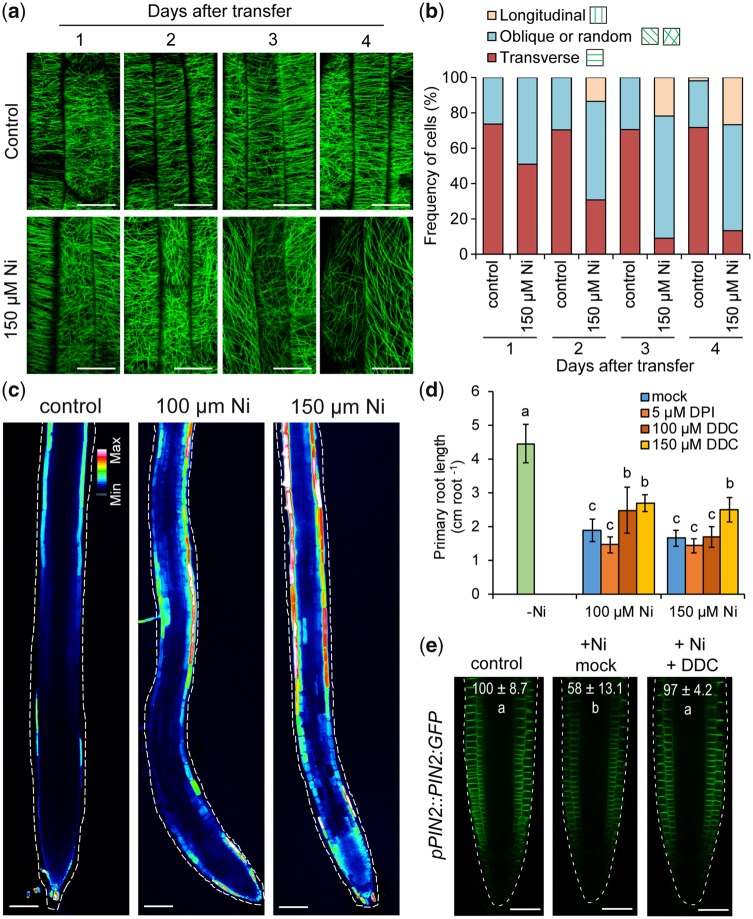
Contamination of soils with heavy metals, such as nickel (Ni), is a major environmental concern due to increasing pollution from industrial activities, burning of fossil fuels, incorrect disposal of sewage sludge, excessive manure application and the use of fertilizers and pesticides in agriculture. Excess Ni induces leaf chlorosis and inhibits plant growth, but the mechanisms underlying growth inhibition remain largely unknown. A detailed analysis of root development in Arabidopsis thaliana in the presence of Ni revealed that this heavy metal induces gravitropic defects and locally inhibits root growth by suppressing cell elongation without significantly disrupting the integrity of the stem cell niche. The analysis of auxin-responsive reporters revealed that excess Ni inhibits shootward auxin distribution. Furthermore, we found that PIN2 is very sensitive to Ni, as the presence of this heavy metal rapidly reduced PIN2 levels in roots. A transcriptome analysis also showed that Ni affects the expression of many genes associated with plant cell walls and that Ni-induced transcriptional changes are largely independent of iron (Fe). In addition, we raised evidence that excess Ni increases the accumulation of reactive oxygen species and disturbs the integrity and orientation of microtubules. Together, our results highlight which processes are primarily targeted by Ni to alter root growth and development.
Keywords: Arabidopsis thaliana; Heavy metals; Polar auxin transport; Reactive oxygen species; Root development; Transcriptome.
� The Author(s) 2019. Published by Oxford University Press on behalf of Japanese Society of Plant Physiologists.
Figures
Comment in
-
Understanding Nickel Responses in Plants: More than Just an Interaction with Iron Homeostasis.Plant Cell Physiol. 2020 Mar 1;61(3):443-444. doi: 10.1093/pcp/pcaa016. Plant Cell Physiol. 2020. PMID: 32142133 No abstract available.
References
-
- Alexa A., Rahnenfuhrer J. (2019) topGO: Enrichment Analysis for Gene Ontology. R Package Version 2.36.0.
-
- Ambrose C., Ruan Y., Gardiner J., Tamblyn L.M., Catching A., Kirik V. (2013) CLASP interacts with Sorting Nexin 1 to link microtubules and auxin transport via PIN2 recycling in Arabidopsis thaliana. Dev. Cell 24: 649–659. - PubMed
-
- Baskin T.I. (2005) Anisotropic expansion of the plant cell wall. Annu. Rev. Cell Dev. Biol. 21: 203–222. - PubMed
-
- Becher M., Talke I.N., Krall L., Kr�mer U. (2004) Cross-species microarray transcript profiling reveals high constitutive expression of metal homeostasis genes in shoots of the zinc hyperaccumulator Arabidopsis halleri. Plant J. 37: 251–268. - PubMed
-
- Beneš I., Schreiber K., Ripperger H., Kircheiss A. (1983) Metal complex formation by nicotianamine, a possible phytosiderophore. Experientia 39: 261–262.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
