

Stiffness Sensing by Cells
- PMID: 31751165
- PMCID: PMC7276923
- DOI: 10.1152/physrev.00013.2019
Stiffness Sensing by Cells
Abstract
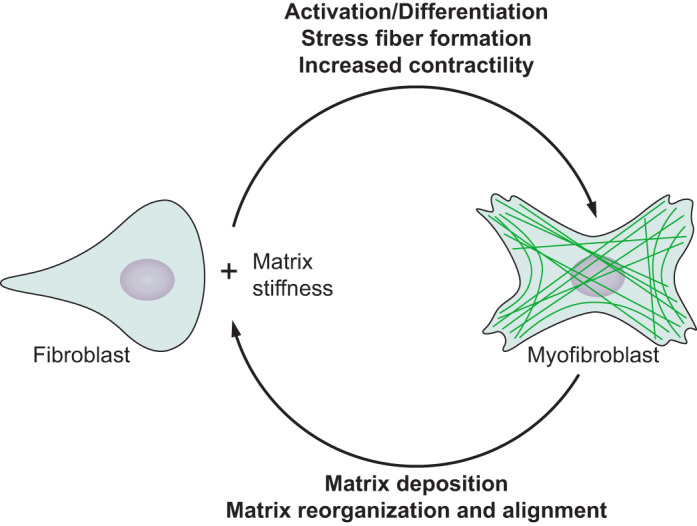
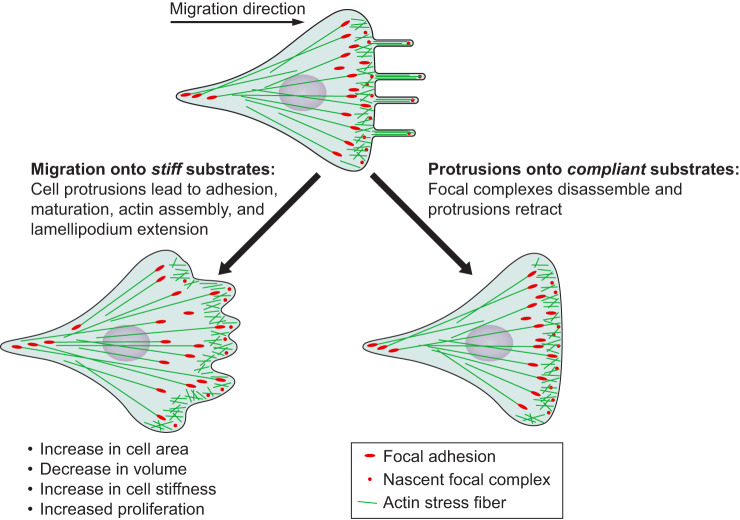
Physical stimuli are essential for the function of eukaryotic cells, and changes in physical signals are important elements in normal tissue development as well as in disease initiation and progression. The complexity of physical stimuli and the cellular signals they initiate are as complex as those triggered by chemical signals. One of the most important, and the focus of this review, is the effect of substrate mechanical properties on cell structure and function. The past decade has produced a nearly exponentially increasing number of mechanobiological studies to define how substrate stiffness alters cell biology using both purified systems and intact tissues. Here we attempt to identify common features of mechanosensing in different systems while also highlighting the numerous informative exceptions to what in early studies appeared to be simple rules by which cells respond to mechanical stresses.
Keywords: cell mechanics; cytoskeleton; mechanobiology; mechanotransduction; substrate stiffness; tissue mechanics; viscoelasticity.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures
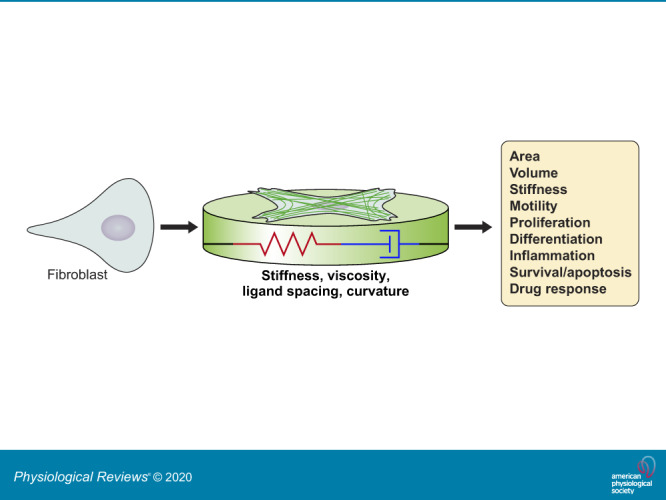
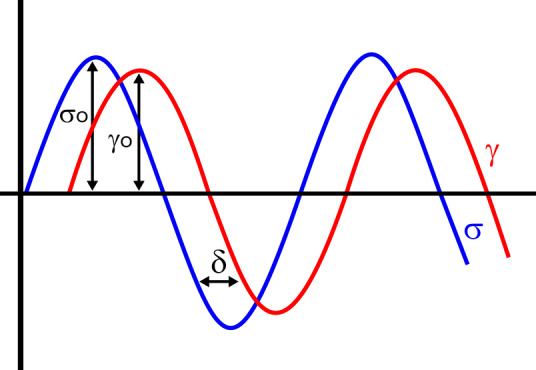
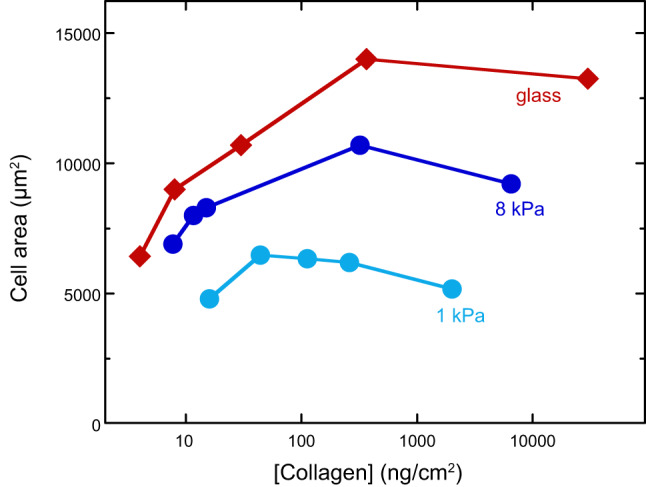
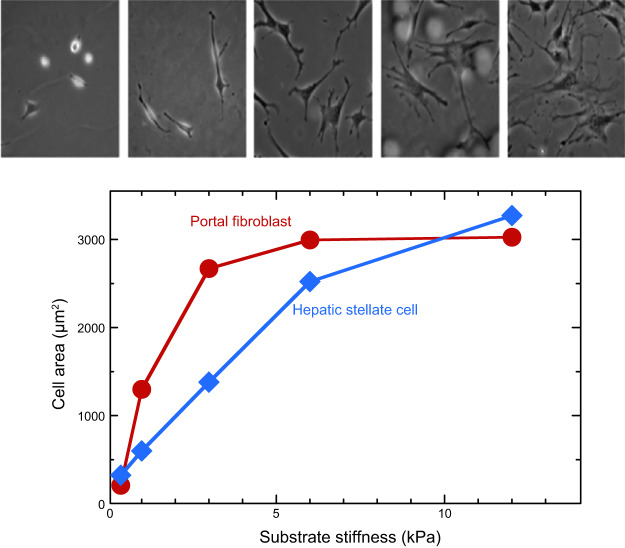
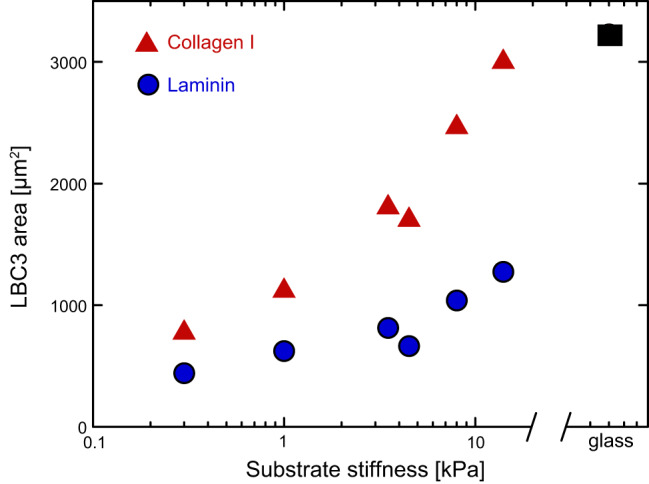
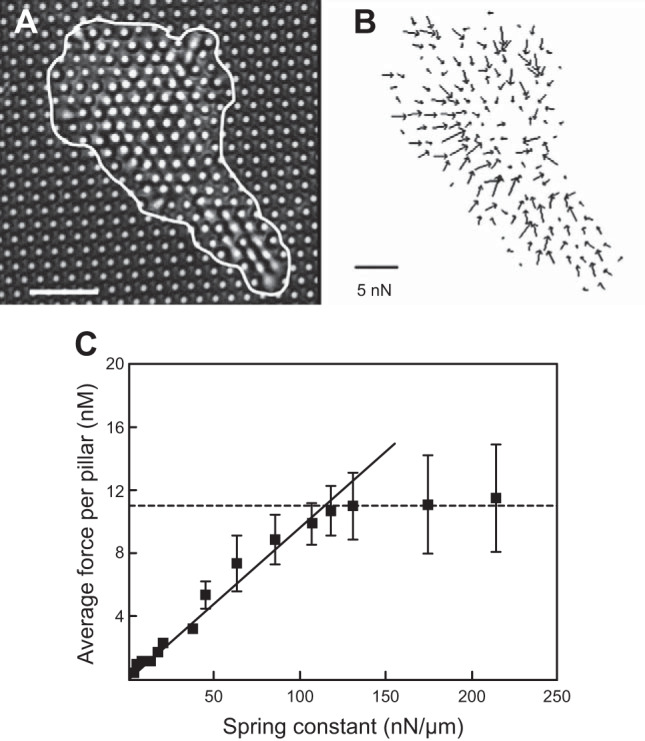
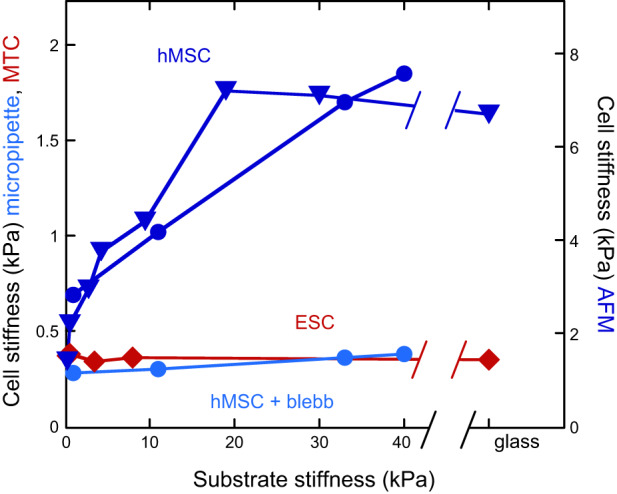
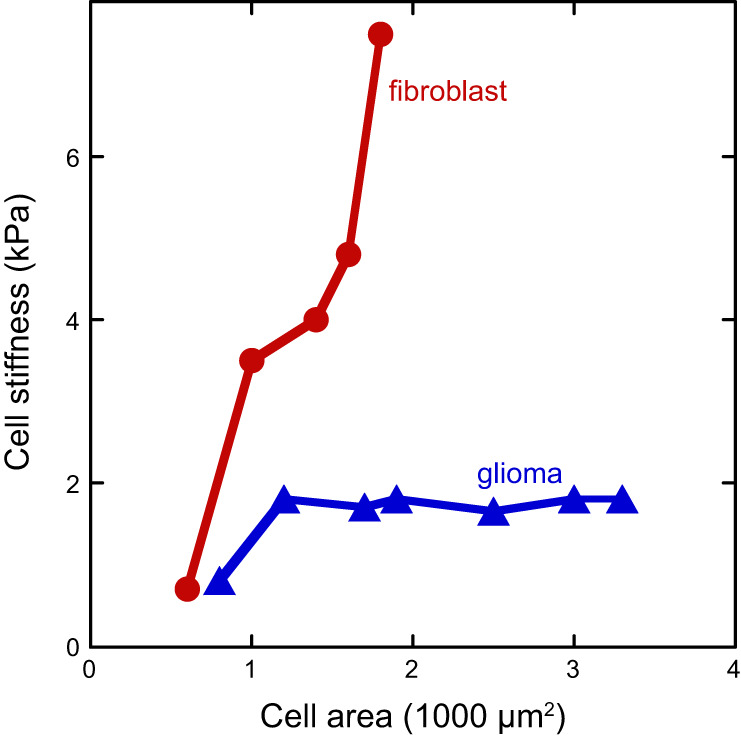
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous