

Recent Advances in EPAC-Targeted Therapies: A Biophysical Perspective
- PMID: 31752286
- PMCID: PMC6912387
- DOI: 10.3390/cells8111462
Recent Advances in EPAC-Targeted Therapies: A Biophysical Perspective
Abstract
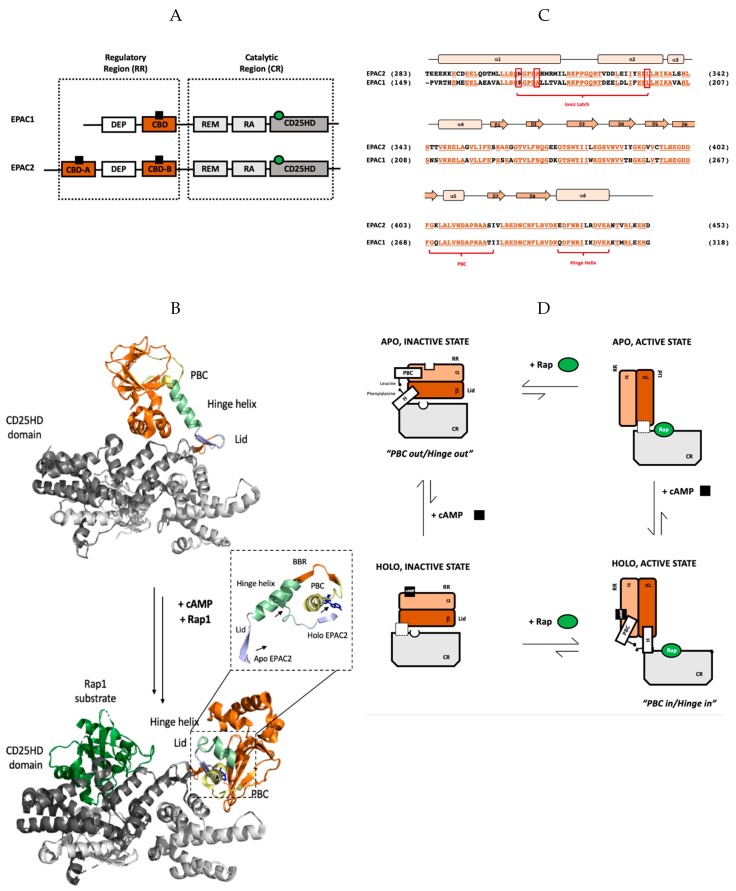
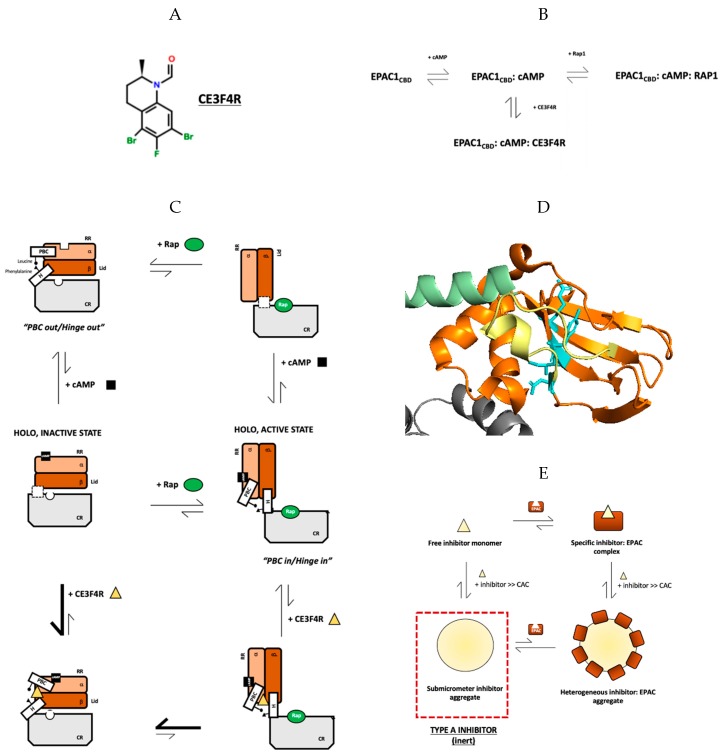
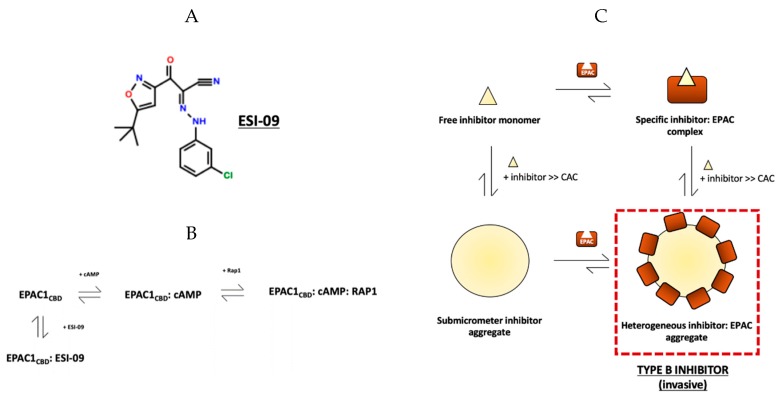
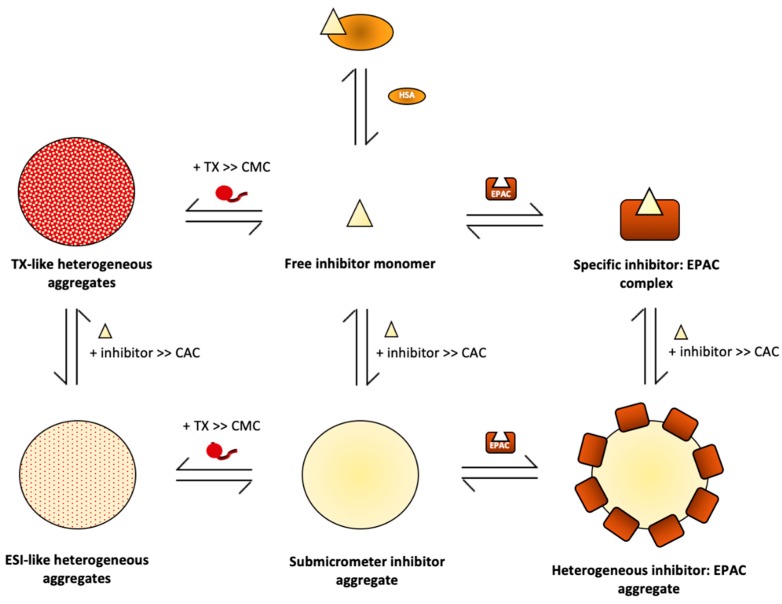
The universal second messenger cAMP regulates diverse intracellular processes by interacting with ubiquitously expressed proteins, such as Protein Kinase A (PKA) and the Exchange Protein directly Activated by cAMP (EPAC). EPAC is implicated in multiple pathologies, thus several EPAC-specific inhibitors have been identified in recent years. However, the mechanisms and molecular interactions underlying the EPAC inhibition elicited by such compounds are still poorly understood. Additionally, being hydrophobic low molecular weight species, EPAC-specific inhibitors are prone to forming colloidal aggregates, which result in non-specific aggregation-based inhibition (ABI) in aqueous systems. Here, we review from a biophysical perspective the molecular basis of the specific and non-specific interactions of two EPAC antagonists-CE3F4R, a non-competitive inhibitor, and ESI-09, a competitive inhibitor of EPAC. Additionally, we discuss the value of common ABI attenuators (e.g., TX and HSA) to reduce false positives at the expense of introducing false negatives when screening aggregation-prone compounds. We hope this review provides the EPAC community effective criteria to evaluate similar compounds, aiding in the optimization of existing drug leads, and informing the development of the next generation of EPAC-specific inhibitors.
Keywords: EPAC; NMR; aggregation-based inhibition; allostery; cAMP; drug design; dynamics; protein-ligand binding; screening; signaling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Berthet J., Rall T.W., Sutherland E.W. The relationship of epinephrine and glucagon to liver phosphorylase. IV. Effect of epinephrine and glucagon on the reactivation of phosphorylase in liver homogenates. J. Biol. Chem. 1957;224:463–475. - PubMed
-
- Ravnskjaer K., Madiraju A., Montminy M. Role of the cAMP pathway in glucose and lipid metabolism. Handb. Exp. Pharmacol. 2015;233:29–49. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
