

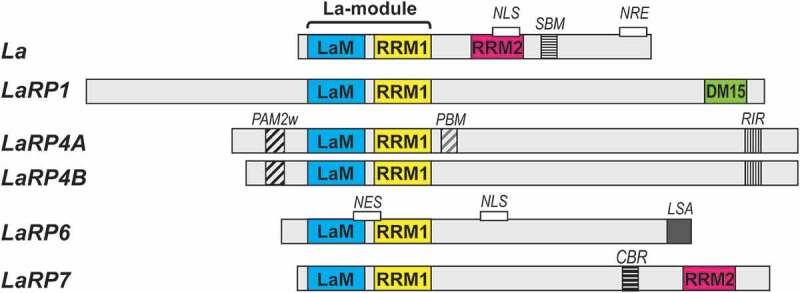
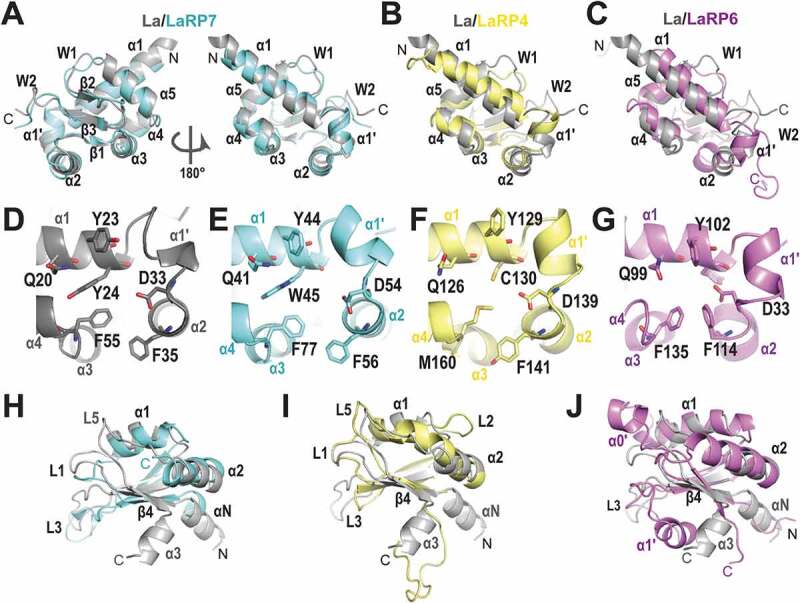
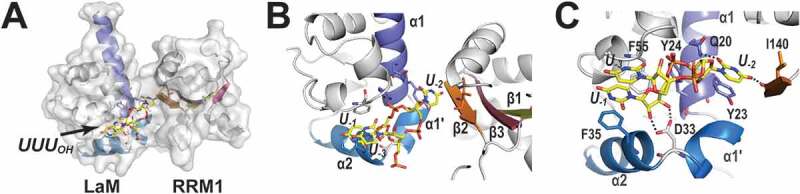
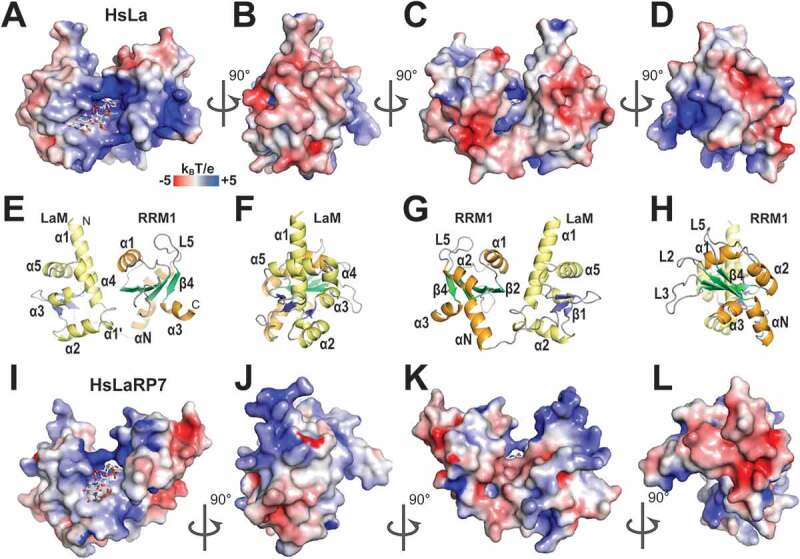
The La-related proteins: structures and interactions of a versatile superfamily of RNA-binding proteins
- PMID: 31752575
- PMCID: PMC7928045
- DOI: 10.1080/15476286.2019.1695712
The La-related proteins: structures and interactions of a versatile superfamily of RNA-binding proteins
Abstract
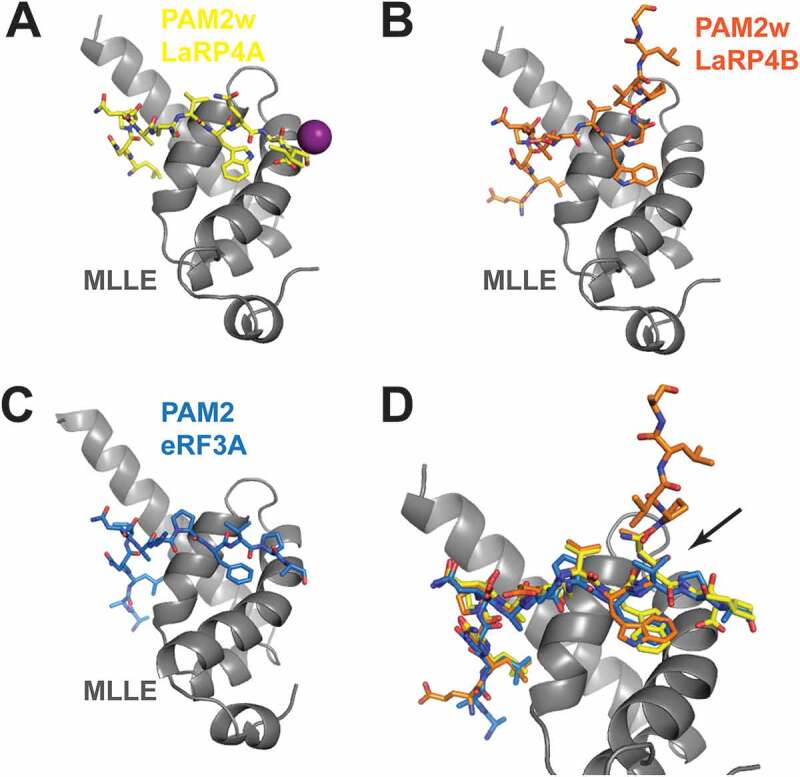
The La-related proteins (LaRPs) are an ancient superfamily of RNA-binding proteins orchestrating the major fates of RNA, from processing and maturation to regulation of mRNA translation. LaRPs are instrumental in modulating complex assemblies where the RNA is bound, folded, processed, escorted and presented to the functional effectors often through recruitment of protein partners. This intricate web of protein-RNA and protein-protein interactions is enabled by the modular nature of the LaRPs, comprising several structured domains connected by flexible linkers, and other sequences lacking recognizable folded motifs. Recent structures, together with biochemical and biophysical studies, have provided insights into how each LaRP family has evolved unique mechanisms of RNA recognition, not only through the conserved RNA-binding unit, the La-module, but also mediated by other family-specific motifs. Furthermore, in a series of unexpected twists and turns, they have revealed that the dynamic and conformational interplay of multi-structured domains and disordered regions operate in unison to achieve RNA substrate discrimination. This review proposes a perspective of our current knowledge of the structure-function relationship of the LaRP superfamily.
Keywords: La-module; La-related proteins; LaRP; RNA biology; RNA-binding proteins; RRM; structure-function relationship.
Conflict of interest statement
No potential conflict of interest was reported by the authors.
Figures
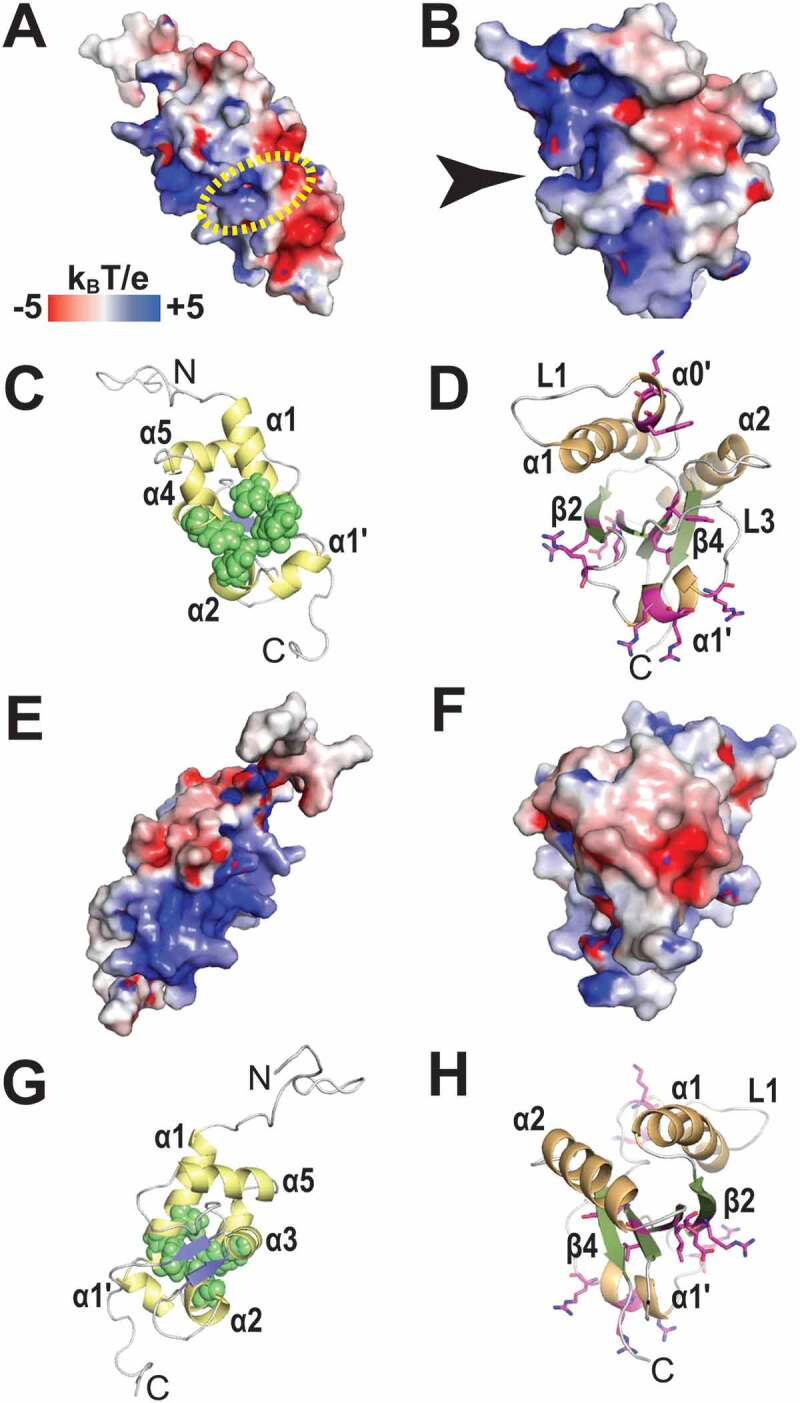
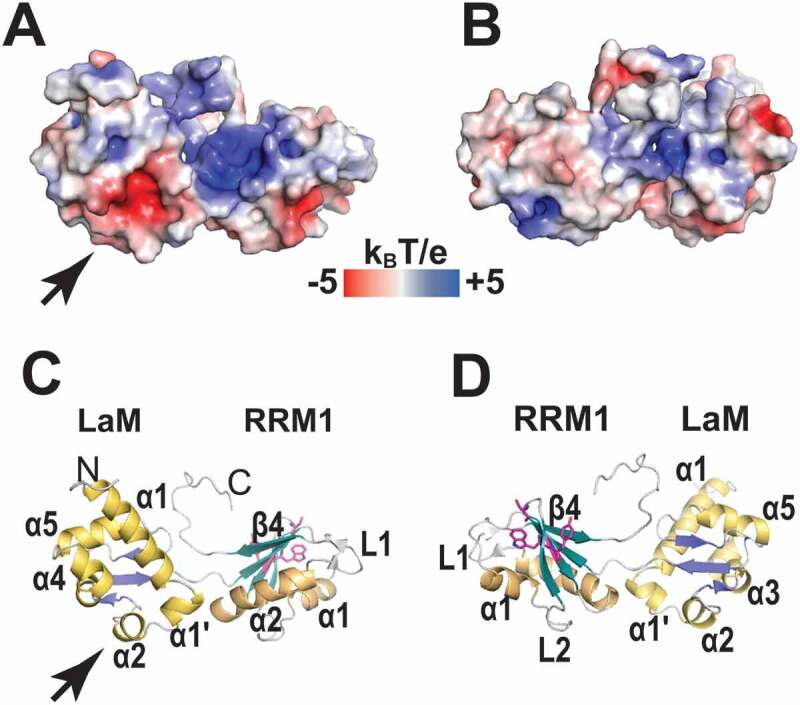
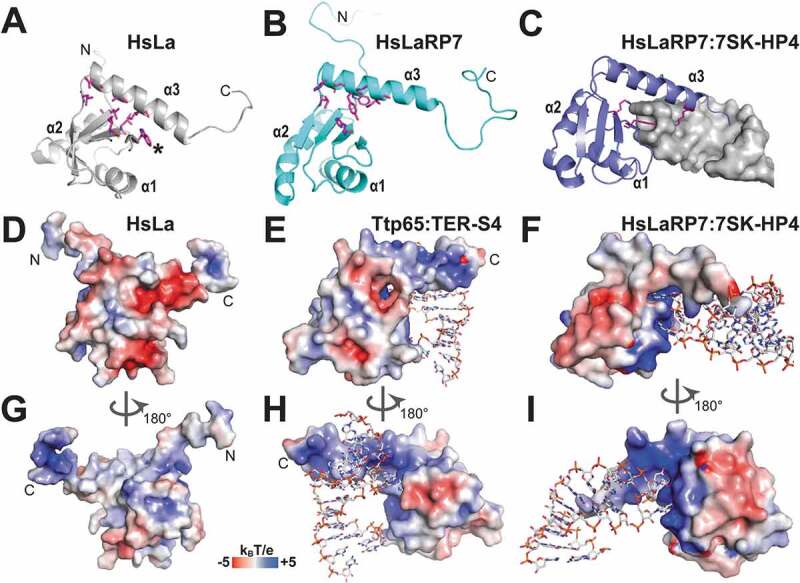
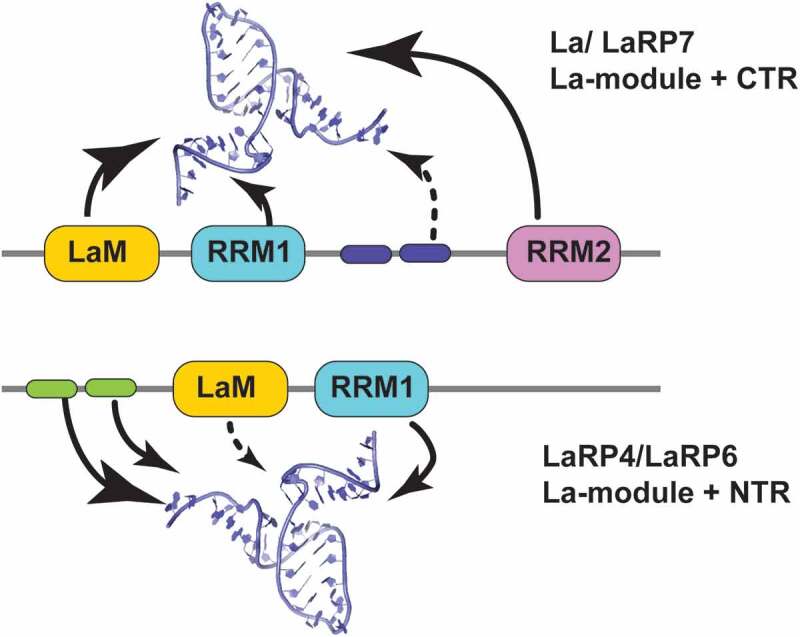
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources