

Protective mechanisms of melatonin against selenium toxicity in Brassica napus: insights into physiological traits, thiol biosynthesis and antioxidant machinery
- PMID: 31752690
- PMCID: PMC6869276
- DOI: 10.1186/s12870-019-2110-6
Protective mechanisms of melatonin against selenium toxicity in Brassica napus: insights into physiological traits, thiol biosynthesis and antioxidant machinery
Abstract
Background: The ubiquitous signaling molecule melatonin (N-acetyl-5-methoxytryptamine) (MT) plays vital roles in plant development and stress tolerance. Selenium (Se) may be phytotoxic at high concentrations. Interactions between MT and Se (IV) stress in higher plants are poorly understood. The aim of this study was to evaluate the defensive roles of exogenous MT (0 μM, 50 μM, and 100 μM) against Se (IV) (0 μM, 50 μM, 100 μM, and 200 μM) stress based on the physiological and biochemical properties, thiol biosynthesis, and antioxidant system of Brassica napus plants subjected to these treatments.
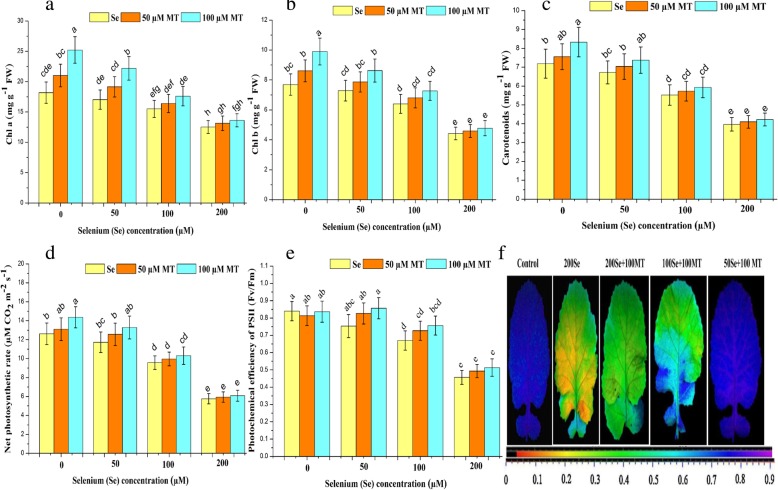
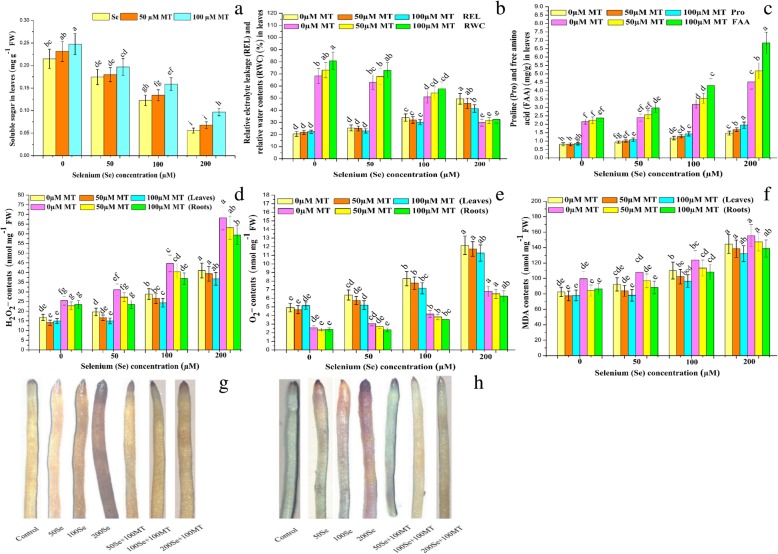
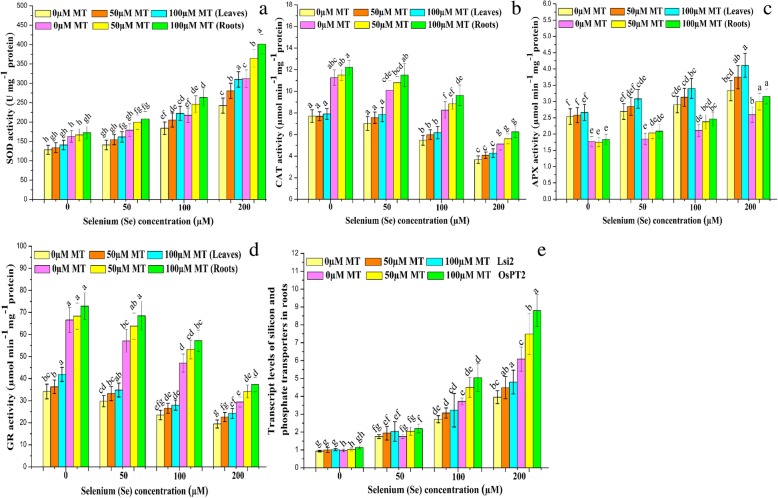
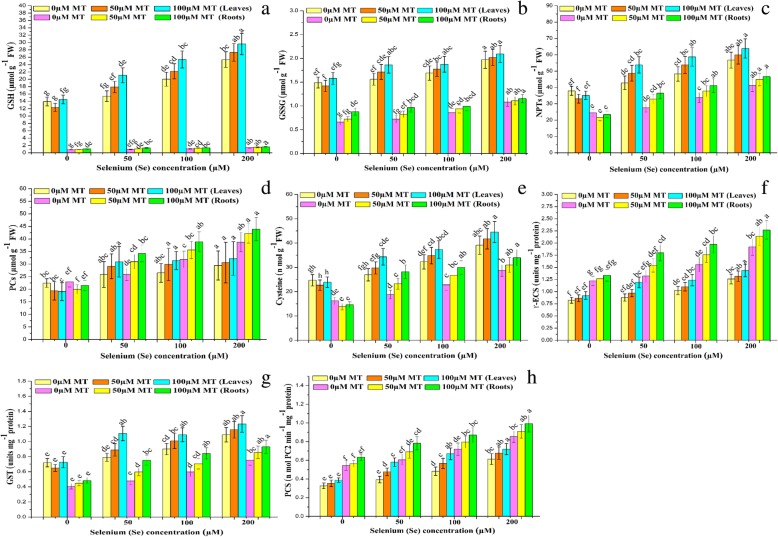
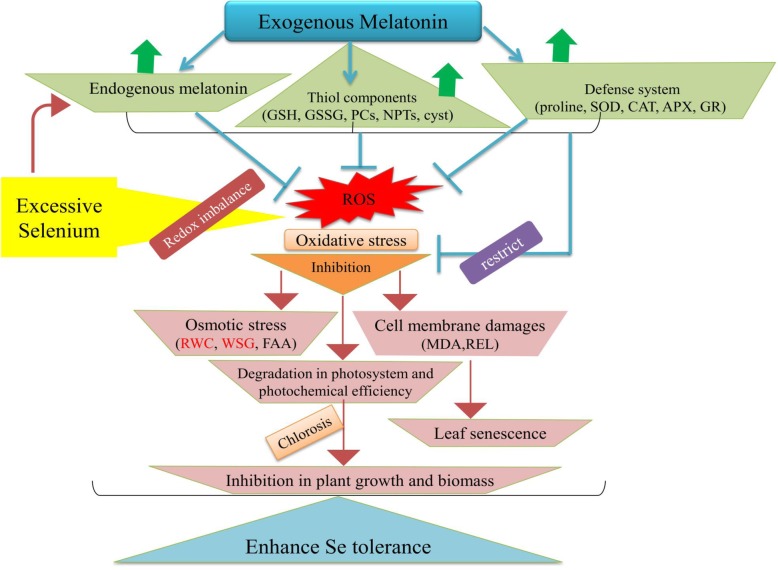
Results: Se (IV) stress inhibited B. napus growth and biomass accumulation, reduced pigment content, and lowered net photosynthetic rate (Pn) and PSII photochemical efficiency (Fv/Fm) in a dose-dependent manner. All of the aforementioned responses were effectively alleviated by exogenous MT treatment. Exogenous MT mitigated oxidative damage and lipid peroxidation and protected the plasma membranes from Se toxicity by reducing Se-induced reactive oxygen species (ROS) accumulation. MT also alleviated osmotic stress by restoring foliar water and sugar levels. Relative to standalone Se treatment, the combination of MT and Se upregulated the ROS-detoxifying enzymes SOD, APX, GR, and CAT, increased proline, free amino acids, and the thiol components GSH, GSSG, GSH/GSSG, NPTs, PCs, and cys and upregulated the metabolic enzymes γ-ECS, GST, and PCS. Therefore, MT application attenuates Se-induce oxidative damage in plants. MT promotes the accumulation of chelating agents in the roots, detoxifies Se there, and impedes its further translocation to the leaves.
Conclusions: Exogenous MT improves the physiological traits, antioxidant system, and thiol ligand biosynthesis in B. napus subjected to Se stress primarily by enhancing Se detoxification and sequestration especially at the root level. Our results reveal better understanding of Se-phytotoxicity and Se-stress alleviation by the adequate supply of MT. The mechanisms of MT-induced plant tolerance to Se stress have potential implications in developing novel strategies for safe crop production in Se-rich soils.
Keywords: Antioxidants; Oilseed rape; Osmolytes; Oxidative stress; Plant growth regulator; Selenium; Thiols.
Conflict of interest statement
“All authors declared that they have no competing interest regarding the submission of this article and its probable publication”.
Figures
Similar articles
-
Mitigation effects of exogenous melatonin-selenium nanoparticles on arsenic-induced stress in Brassica napus.Environ Pollut. 2022 Jan 1;292(Pt B):118473. doi: 10.1016/j.envpol.2021.118473. Epub 2021 Nov 7. Environ Pollut. 2022. PMID: 34758366
-
Exogenous melatonin improves the salt tolerance of cotton by removing active oxygen and protecting photosynthetic organs.BMC Plant Biol. 2021 Jul 10;21(1):331. doi: 10.1186/s12870-021-03082-7. BMC Plant Biol. 2021. PMID: 34246235 Free PMC article.
-
Selenium mitigates the chromium toxicity in Brassicca napus L. by ameliorating nutrients uptake, amino acids metabolism and antioxidant defense system.Plant Physiol Biochem. 2019 Dec;145:142-152. doi: 10.1016/j.plaphy.2019.10.035. Epub 2019 Oct 25. Plant Physiol Biochem. 2019. PMID: 31689666
-
Role of selenium toxicity and oxidative stress in aquatic birds.Aquat Toxicol. 2002 Apr;57(1-2):11-26. doi: 10.1016/s0166-445x(01)00263-6. Aquat Toxicol. 2002. PMID: 11879935 Review.
-
Using Exogenous Melatonin, Glutathione, Proline, and Glycine Betaine Treatments to Combat Abiotic Stresses in Crops.Int J Mol Sci. 2022 Oct 26;23(21):12913. doi: 10.3390/ijms232112913. Int J Mol Sci. 2022. PMID: 36361700 Free PMC article. Review.
Cited by
-
Selenium seed priming enhanced the growth of salt-stressed Brassica rapa L. through improving plant nutrition and the antioxidant system.Front Plant Sci. 2023 Jan 13;13:1050359. doi: 10.3389/fpls.2022.1050359. eCollection 2022. Front Plant Sci. 2023. PMID: 36714767 Free PMC article.
-
Melatonin improves the germination rate of cotton seeds under drought stress by opening pores in the seed coat.PeerJ. 2020 Jul 6;8:e9450. doi: 10.7717/peerj.9450. eCollection 2020. PeerJ. 2020. PMID: 32704446 Free PMC article.
-
ROS and NO Phytomelatonin-Induced Signaling Mechanisms under Metal Toxicity in Plants: A Review.Antioxidants (Basel). 2021 May 13;10(5):775. doi: 10.3390/antiox10050775. Antioxidants (Basel). 2021. PMID: 34068211 Free PMC article. Review.
-
Exogenous melatonin enhances low-temperature stress of jute seedlings through modulation of photosynthesis and antioxidant potential.Heliyon. 2023 Aug 14;9(8):e19125. doi: 10.1016/j.heliyon.2023.e19125. eCollection 2023 Aug. Heliyon. 2023. PMID: 37636352 Free PMC article.
-
Transcriptome analysis reveals the promoting effects of exogenous melatonin on the selenium uptake in grape under selenium stress.Front Plant Sci. 2024 Aug 22;15:1447451. doi: 10.3389/fpls.2024.1447451. eCollection 2024. Front Plant Sci. 2024. PMID: 39239199 Free PMC article.
References
-
- Drahonovský J, Szkova J, Mestek O, Tremlova J, Kana A, Najmanova J, Tlustos P. Selenium uptake, transformation and inter-element interactions by selected wildlife plant species after foliar selenate application. Environ Exp Bot. 2016;125:12–19. doi: 10.1016/j.envexpbot.2016.01.006. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
