

Autophagy and cancer: Modulation of cell death pathways and cancer cell adaptations
- PMID: 31753861
- PMCID: PMC7039213
- DOI: 10.1083/jcb.201909033
Autophagy and cancer: Modulation of cell death pathways and cancer cell adaptations
Abstract
Autophagy is intricately linked with many intracellular signaling pathways, particularly nutrient-sensing mechanisms and cell death signaling cascades. In cancer, the roles of autophagy are context dependent. Tumor cell-intrinsic effects of autophagy can be both tumor suppressive and tumor promotional. Autophagy can therefore not only activate and inhibit cell death, but also facilitate the switch between cell death mechanisms. Moreover, autophagy can play opposing roles in the tumor microenvironment via non-cell-autonomous mechanisms. Preclinical data support a tumor-promotional role of autophagy in established tumors and during cancer therapy; this has led to the launch of dozens of clinical trials targeting autophagy in multiple cancer types. However, many questions remain: which tumors and genetic backgrounds are the most sensitive to autophagy inhibition, and which therapies should be combined with autophagy inhibitors? Additionally, since cancer cells are under selective pressure and are prone to adaptation, particularly after treatment, it is unclear if and how cells adapt to autophagy inhibition. Here we review recent literature addressing these issues.
© 2019 Towers et al.
Figures
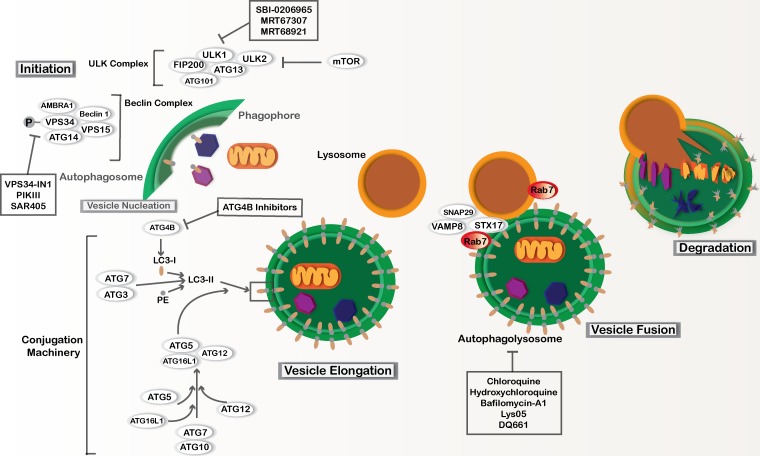
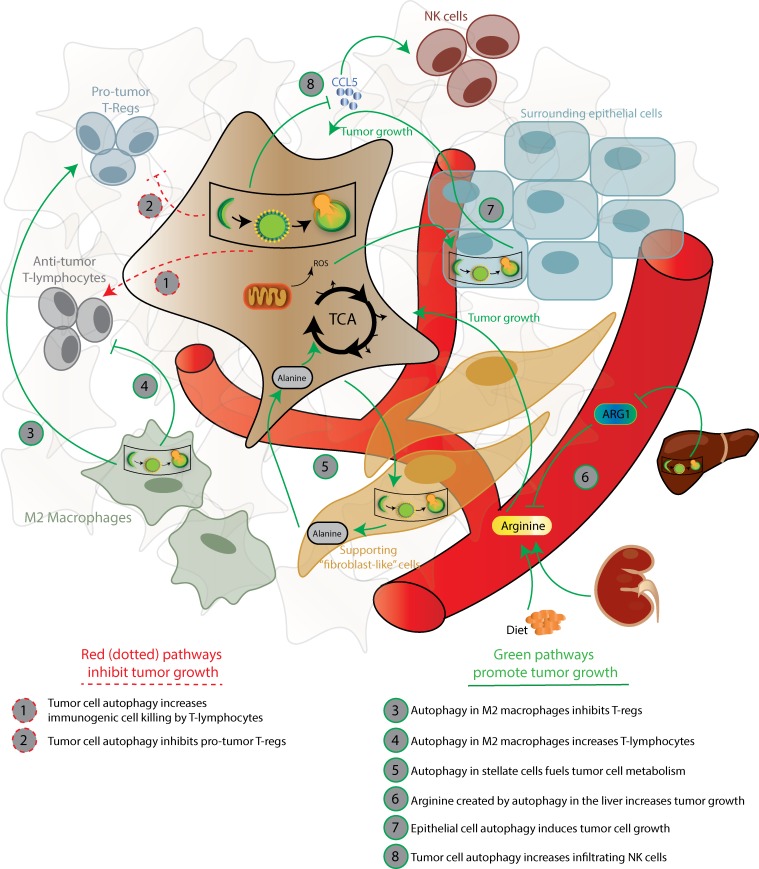
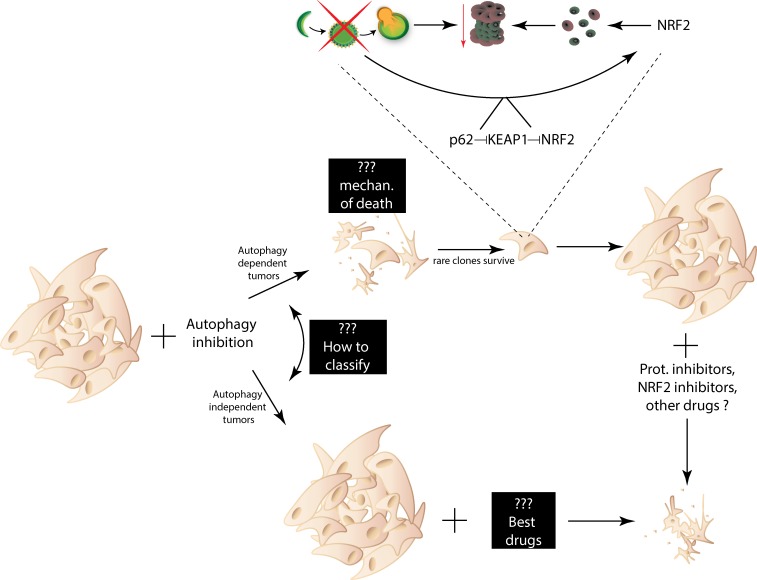
References
-
- Baginska J., Viry E., Berchem G., Poli A., Noman M.Z., van Moer K., Medves S., Zimmer J., Oudin A., Niclou S.P., et al. . 2013. Granzyme B degradation by autophagy decreases tumor cell susceptibility to natural killer-mediated lysis under hypoxia. Proc. Natl. Acad. Sci. USA. 110:17450–17455. 10.1073/pnas.1304790110 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
