

Computational design of a modular protein sense-response system
- PMID: 31754004
- PMCID: PMC7343396
- DOI: 10.1126/science.aax8780
Computational design of a modular protein sense-response system
Abstract
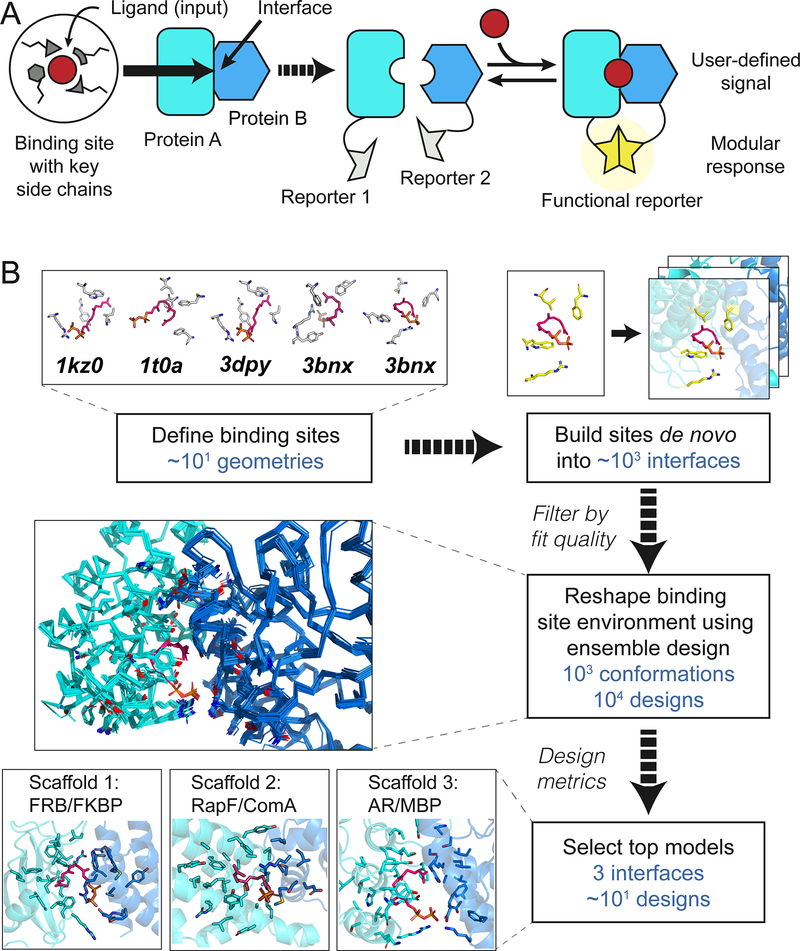
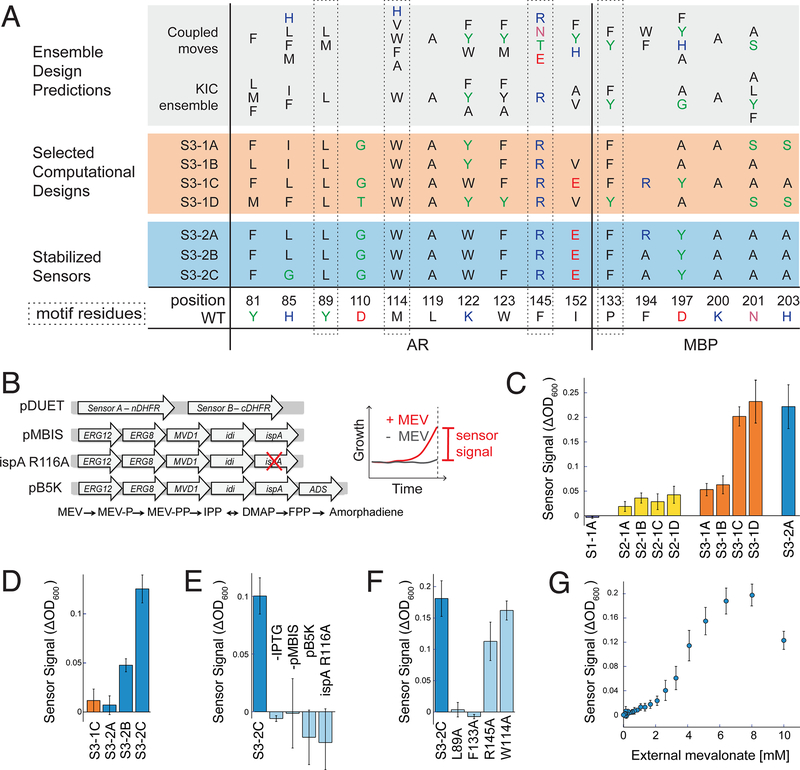
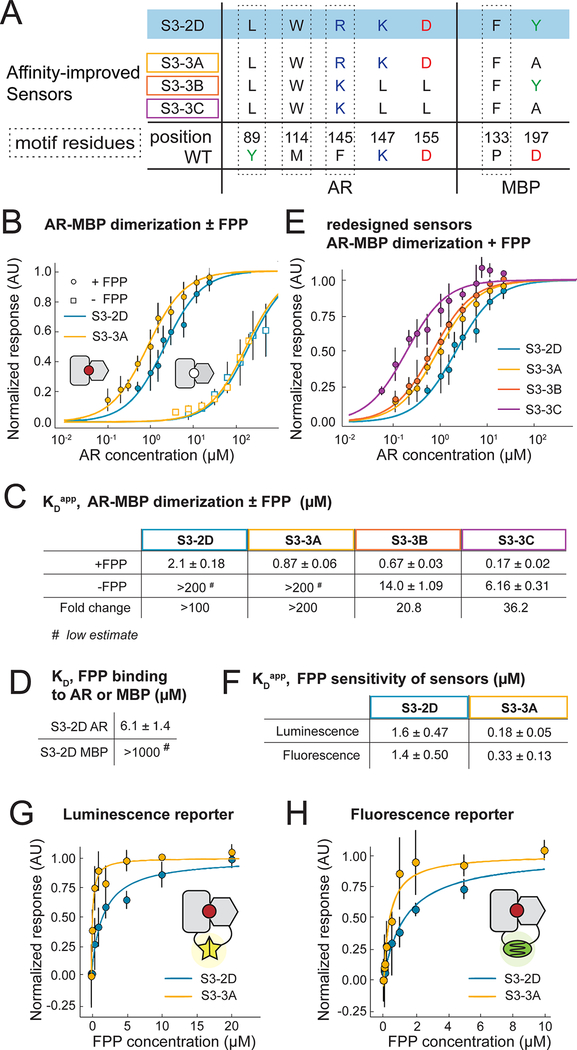
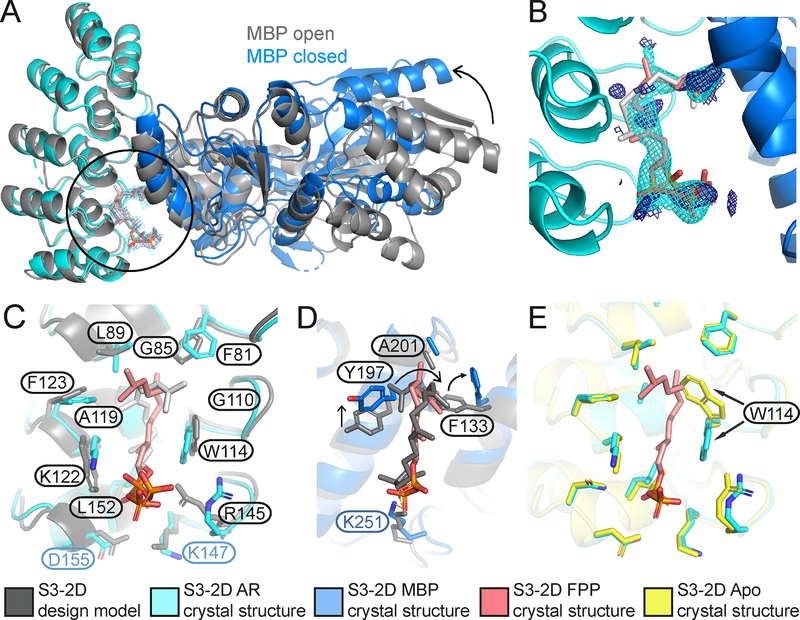
Sensing and responding to signals is a fundamental ability of living systems, but despite substantial progress in the computational design of new protein structures, there is no general approach for engineering arbitrary new protein sensors. Here, we describe a generalizable computational strategy for designing sensor-actuator proteins by building binding sites de novo into heterodimeric protein-protein interfaces and coupling ligand sensing to modular actuation through split reporters. Using this approach, we designed protein sensors that respond to farnesyl pyrophosphate, a metabolic intermediate in the production of valuable compounds. The sensors are functional in vitro and in cells, and the crystal structure of the engineered binding site closely matches the design model. Our computational design strategy opens broad avenues to link biological outputs to new signals.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Designer sense-response systems.Science. 2019 Nov 22;366(6468):952-953. doi: 10.1126/science.aaz8085. Science. 2019. PMID: 31753985 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
