

Mammalian germ cells are determined after PGC colonization of the nascent gonad
- PMID: 31754036
- PMCID: PMC6925976
- DOI: 10.1073/pnas.1910733116
Mammalian germ cells are determined after PGC colonization of the nascent gonad
Abstract
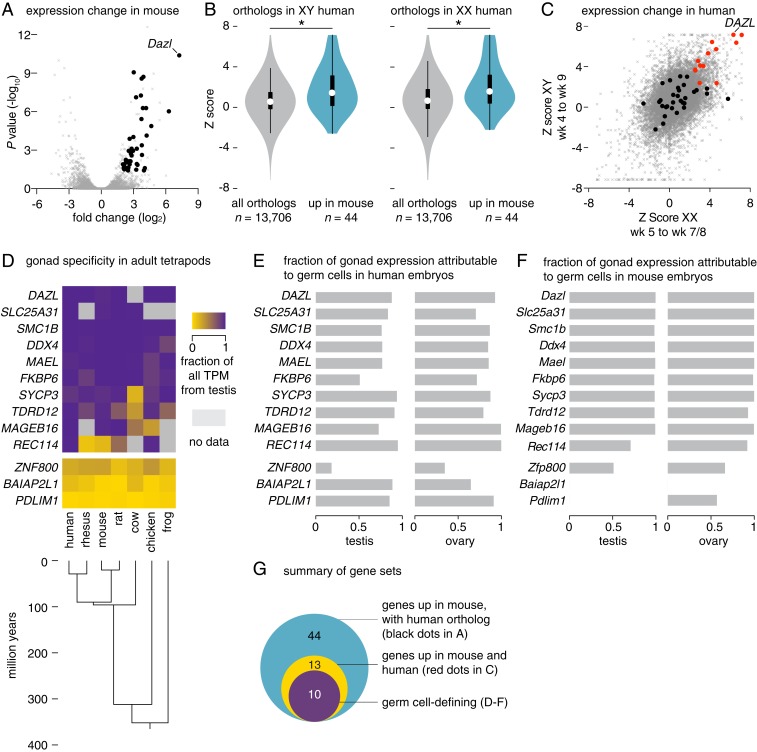
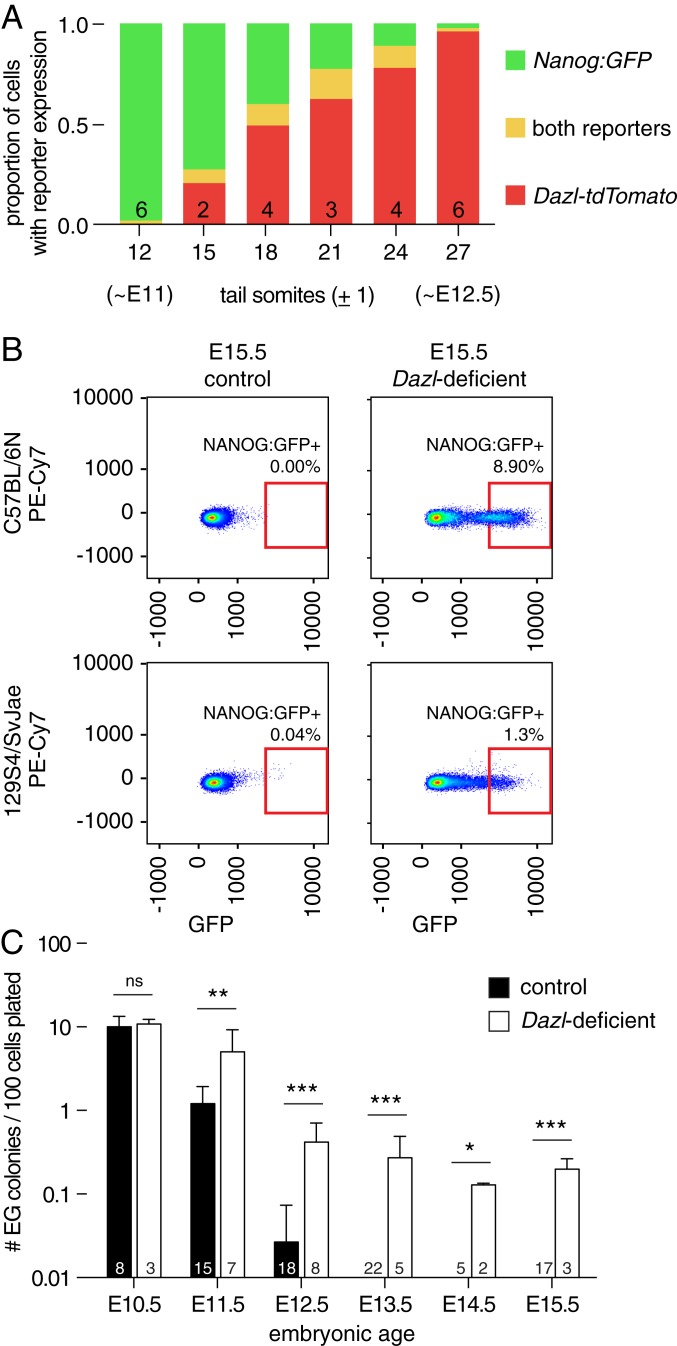
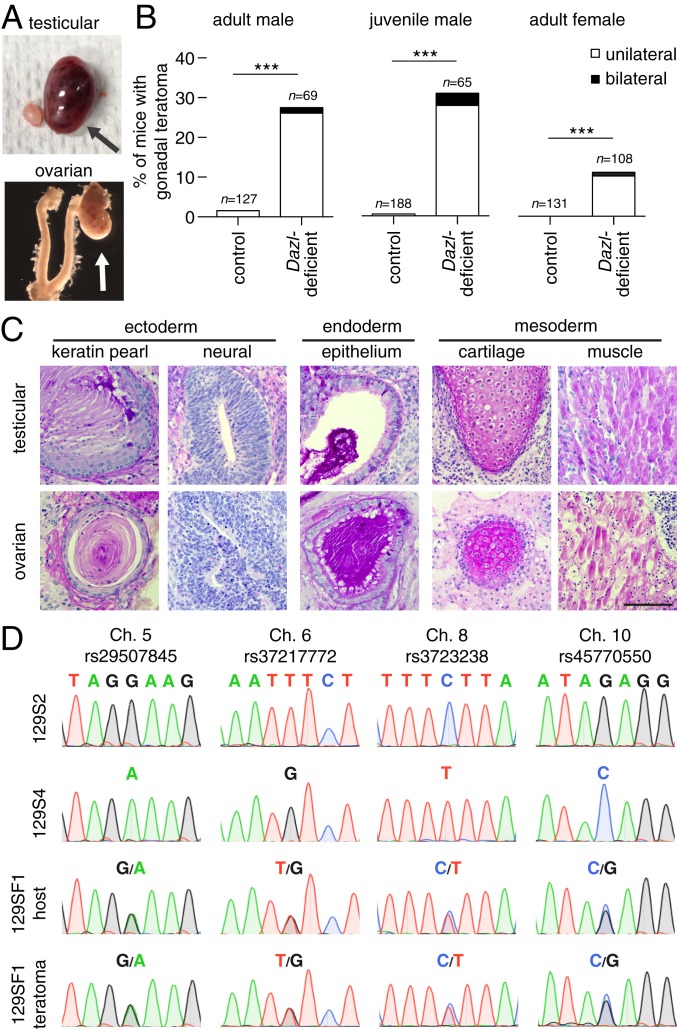
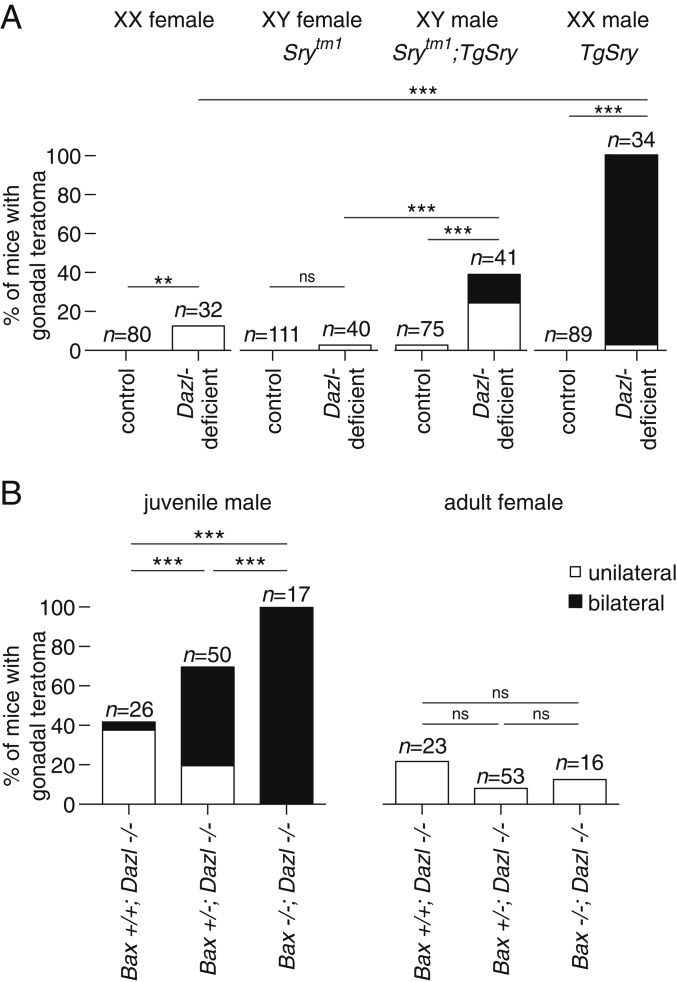
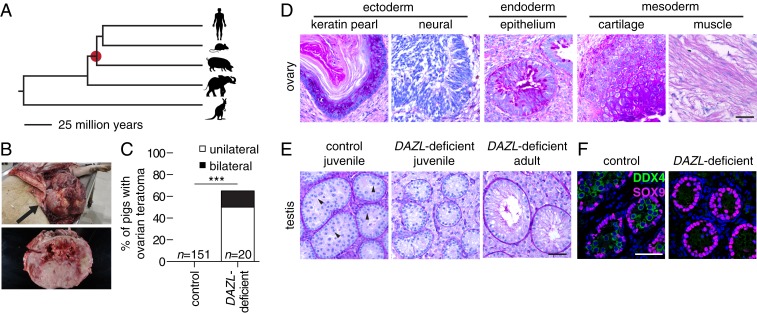
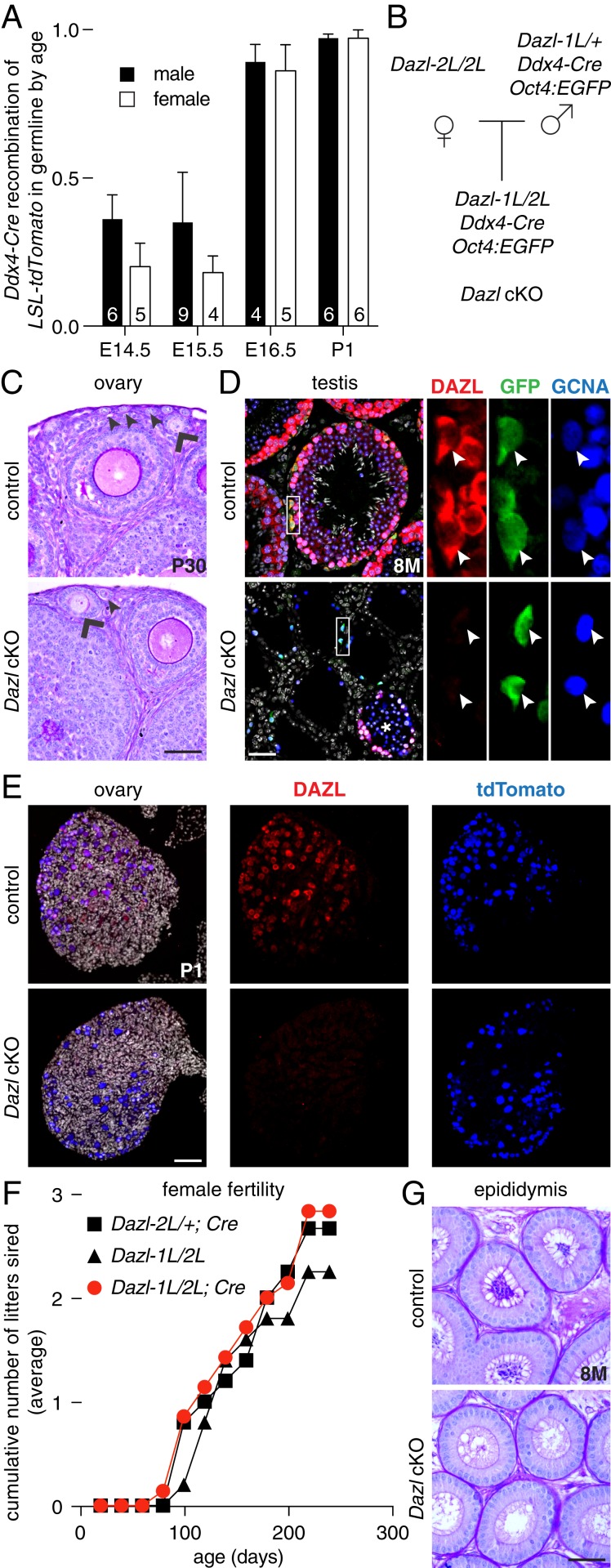
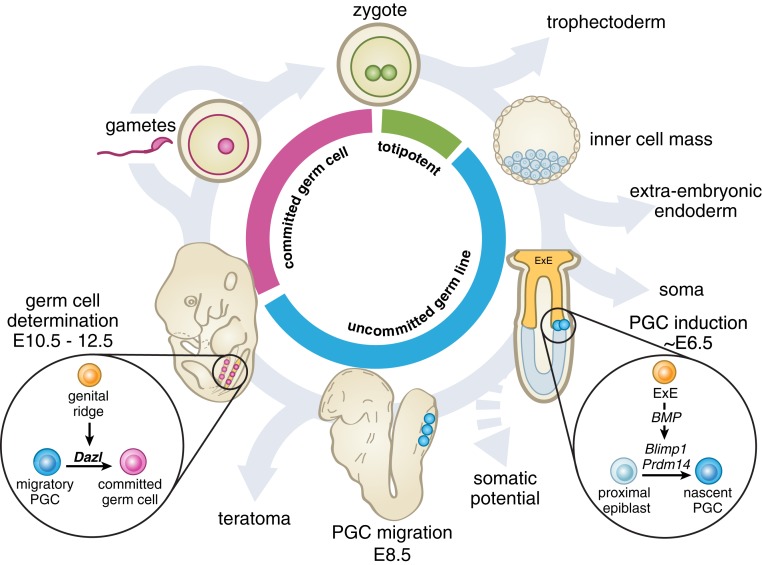
Mammalian primordial germ cells (PGCs) are induced in the embryonic epiblast, before migrating to the nascent gonads. In fish, frogs, and birds, the germline segregates even earlier, through the action of maternally inherited germ plasm. Across vertebrates, migrating PGCs retain a broad developmental potential, regardless of whether they were induced or maternally segregated. In mammals, this potential is indicated by expression of pluripotency factors, and the ability to generate teratomas and pluripotent cell lines. How the germline loses this developmental potential remains unknown. Our genome-wide analyses of embryonic human and mouse germlines reveal a conserved transcriptional program, initiated in PGCs after gonadal colonization, that differentiates germ cells from their germline precursors and from somatic lineages. Through genetic studies in mice and pigs, we demonstrate that one such gonad-induced factor, the RNA-binding protein DAZL, is necessary in vivo to restrict the developmental potential of the germline; DAZL's absence prolongs expression of a Nanog pluripotency reporter, facilitates derivation of pluripotent cell lines, and causes spontaneous gonadal teratomas. Based on these observations in humans, mice, and pigs, we propose that germ cells are determined after gonadal colonization in mammals. We suggest that germ cell determination was induced late in embryogenesis-after organogenesis has begun-in the common ancestor of all vertebrates, as in modern mammals, where this transition is induced by somatic cells of the gonad. We suggest that failure of this process of germ cell determination likely accounts for the origin of human testis cancer.
Keywords: Dazl; commitment; germ cell; pluripotency; teratoma.
Conflict of interest statement
Competing interest statement: A.L.W., D.F.C., and S.C.F. are employees and shareholders of Recombinetics Inc. I.D. is a member of the scientific advisory board of Recombinetics Inc. The remaining authors have declared that no competing interests exist.
Figures
Comment in
-
Quelling germ cell pluripotency on the genital ridge.Proc Natl Acad Sci U S A. 2019 Dec 17;116(51):25374-25375. doi: 10.1073/pnas.1918899116. Epub 2019 Nov 22. Proc Natl Acad Sci U S A. 2019. PMID: 31757856 Free PMC article. No abstract available.
References
-
- Ohinata Y., et al. , A signaling principle for the specification of the germ cell lineage in mice. Cell 137, 571–584 (2009). - PubMed
-
- Extavour C. G., Akam M., Mechanisms of germ cell specification across the metazoans: Epigenesis and preformation. Development 130, 5869–5884 (2003). - PubMed
-
- Wylie C. C., Heasman J., Snape A., O’Driscoll M., Holwill S., Primordial germ cells of Xenopus laevis are not irreversibly determined early in development. Dev. Biol. 112, 66–72 (1985).
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
