

Multiple roles of DNA2 nuclease/helicase in DNA metabolism, genome stability and human diseases
- PMID: 31754720
- PMCID: PMC6943134
- DOI: 10.1093/nar/gkz1101
Multiple roles of DNA2 nuclease/helicase in DNA metabolism, genome stability and human diseases
Abstract
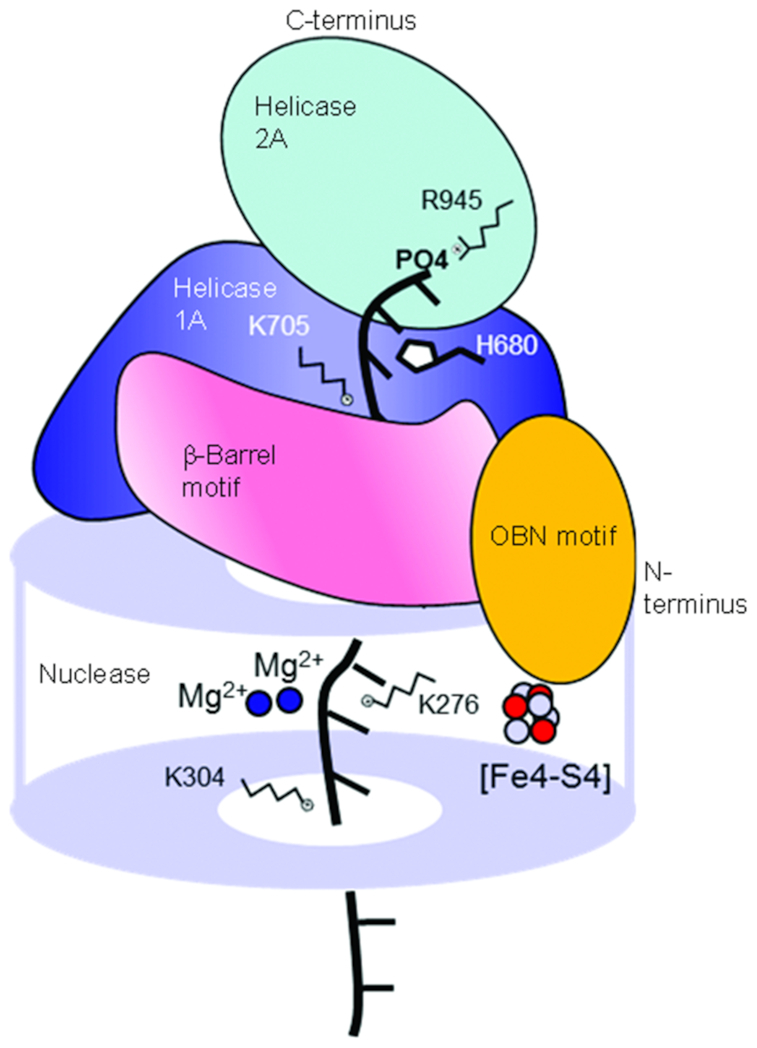
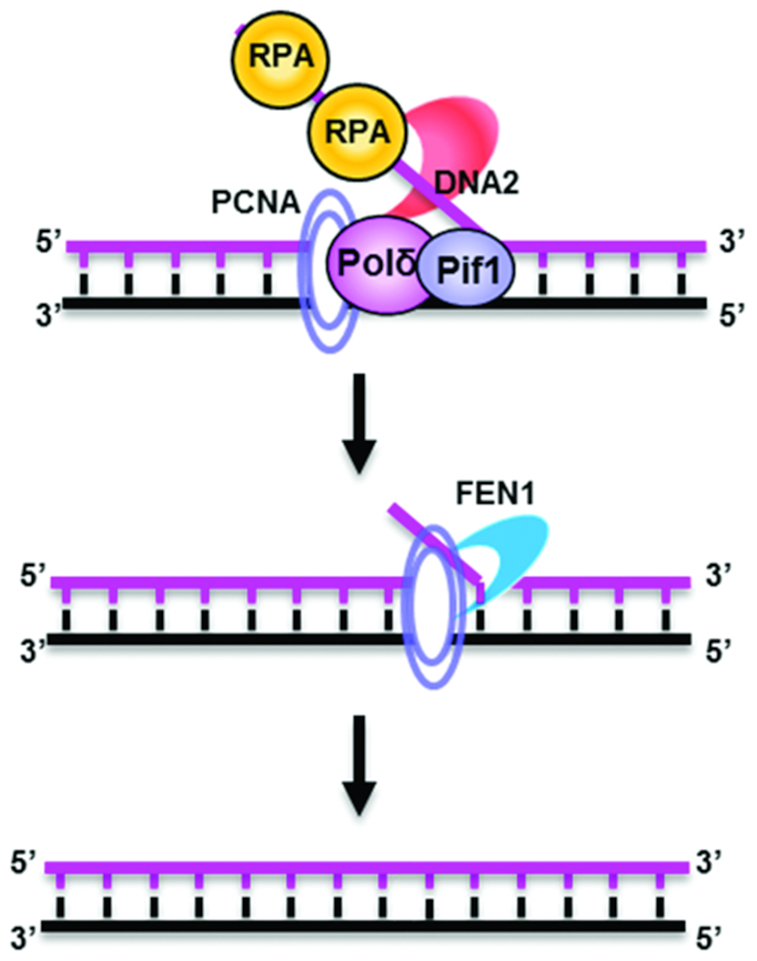
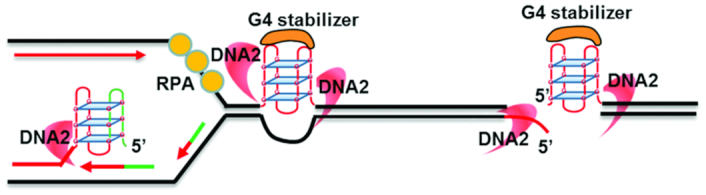
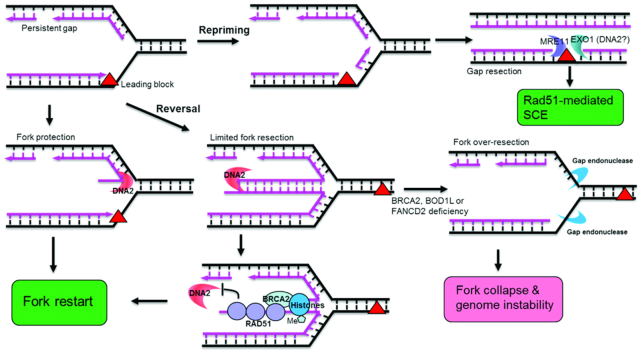
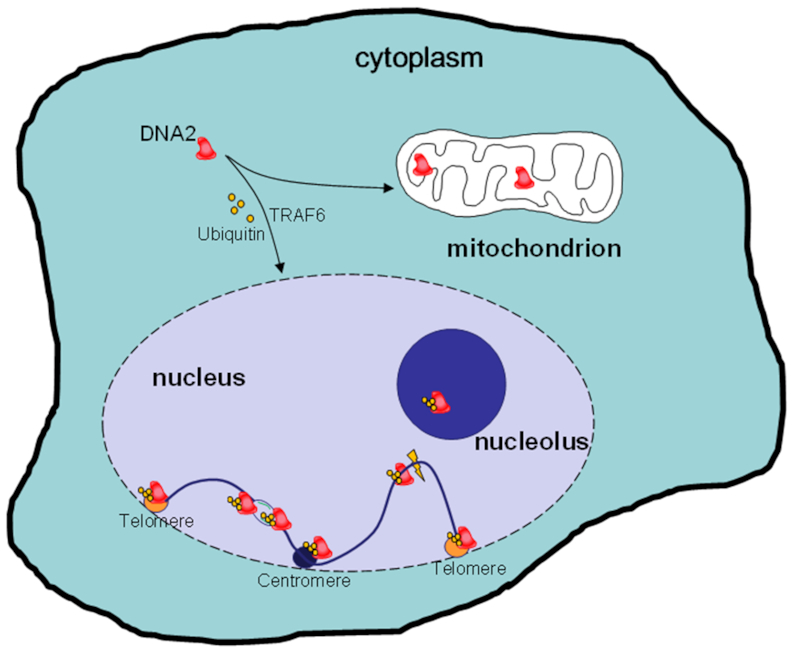
DNA2 nuclease/helicase is a structure-specific nuclease, 5'-to-3' helicase, and DNA-dependent ATPase. It is involved in multiple DNA metabolic pathways, including Okazaki fragment maturation, replication of 'difficult-to-replicate' DNA regions, end resection, stalled replication fork processing, and mitochondrial genome maintenance. The participation of DNA2 in these different pathways is regulated by its interactions with distinct groups of DNA replication and repair proteins and by post-translational modifications. These regulatory mechanisms induce its recruitment to specific DNA replication or repair complexes, such as DNA replication and end resection machinery, and stimulate its efficient cleavage of various structures, for example, to remove RNA primers or to produce 3' overhangs at telomeres or double-strand breaks. Through these versatile activities at replication forks and DNA damage sites, DNA2 functions as both a tumor suppressor and promoter. In normal cells, it suppresses tumorigenesis by maintaining the genomic integrity. Thus, DNA2 mutations or functional deficiency may lead to cancer initiation. However, DNA2 may also function as a tumor promoter, supporting cancer cell survival by counteracting replication stress. Therefore, it may serve as an ideal target to sensitize advanced DNA2-overexpressing cancers to current chemo- and radiotherapy regimens.
© The Author(s) 2019. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
