

Resolving noise-control conflict by gene duplication
- PMID: 31756183
- PMCID: PMC6874299
- DOI: 10.1371/journal.pbio.3000289
Resolving noise-control conflict by gene duplication
Abstract
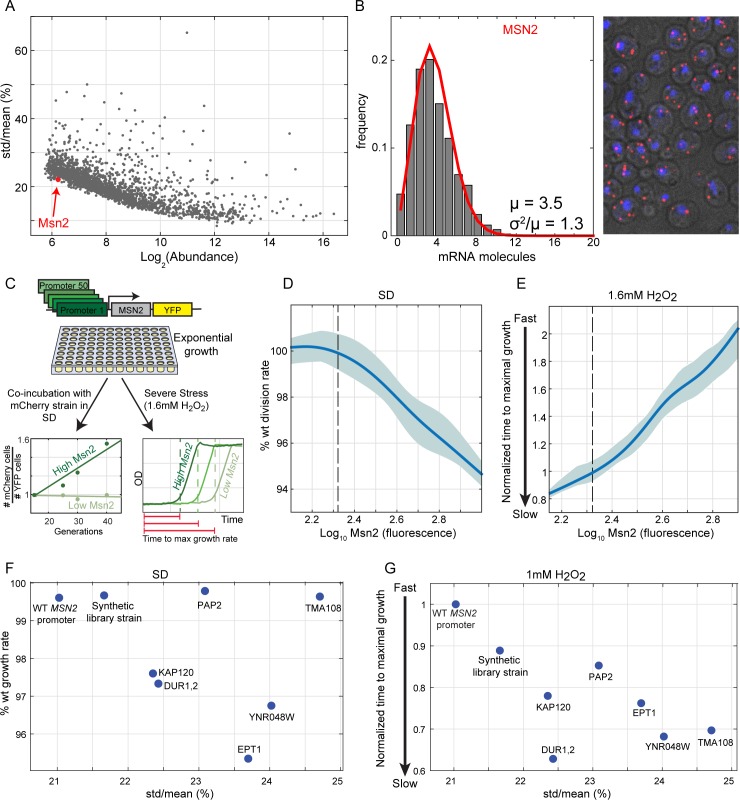
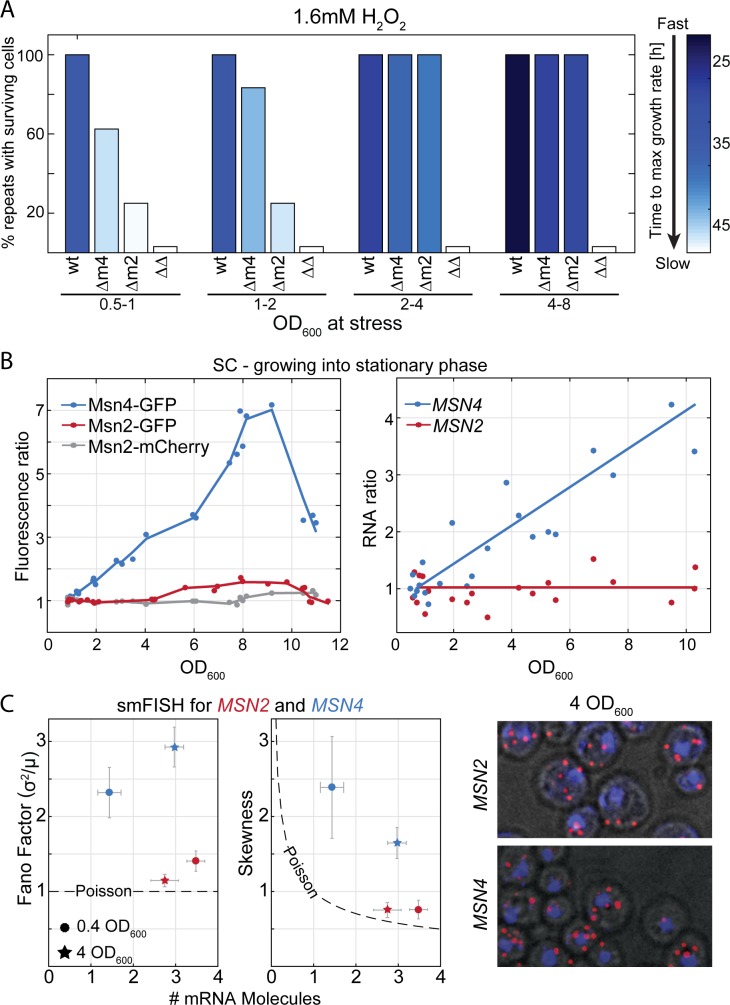
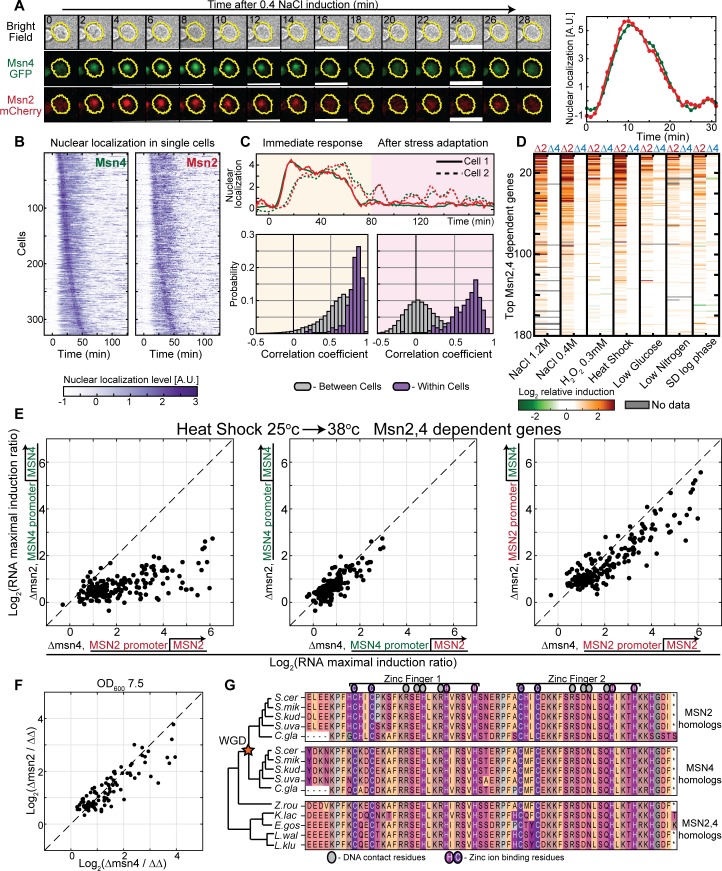
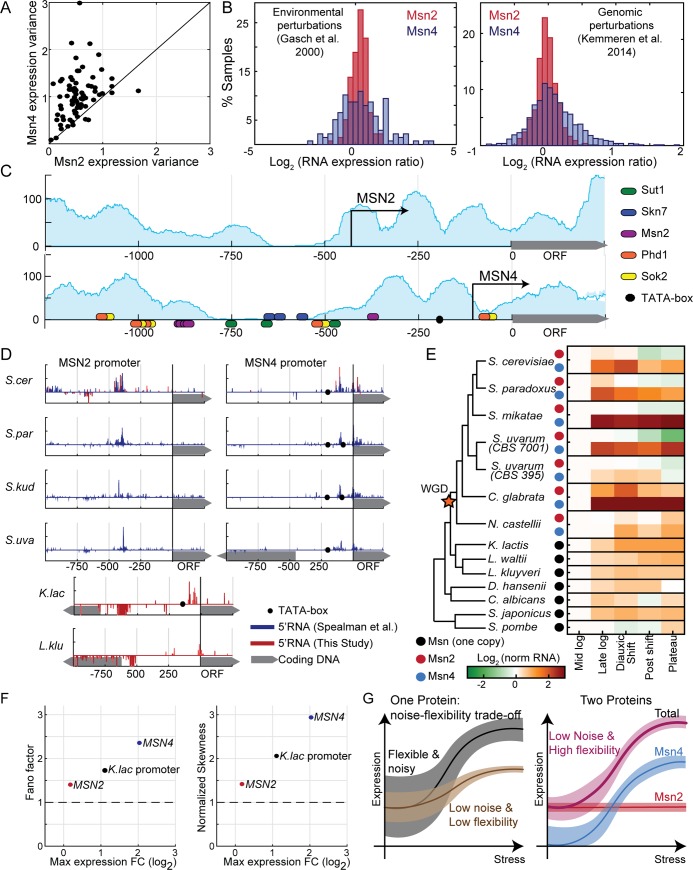
Gene duplication promotes adaptive evolution in two main ways: allowing one duplicate to evolve a new function and splitting ancestral functions between the duplicates. The second scenario may resolve adaptive conflicts that can rise when one gene performs different functions. In an apparent departure from both scenarios, low-expressing transcription factor (TF) duplicates commonly bind to the same DNA motifs and act in overlapping conditions. To examine for possible benefits of this apparent redundancy, we examined the Msn2 and Msn4 duplicates in budding yeast. We show that Msn2,4 function as one unit by inducing the same set of target genes in overlapping conditions. Yet, the two-factor composition allows this unit's expression to be both environmentally responsive and with low noise, resolving an adaptive conflict that limits expression of single genes. We propose that duplication can provide adaptive benefit through cooperation rather than functional divergence, allowing two-factor dynamics with beneficial properties that cannot be achieved by a single gene.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
Regulation plays a multifaceted role in the retention of gene duplicates.PLoS Biol. 2019 Nov 22;17(11):e3000519. doi: 10.1371/journal.pbio.3000519. eCollection 2019 Nov. PLoS Biol. 2019. PMID: 31756186 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
