

Influenza H7N9 Virus Neuraminidase-Specific Human Monoclonal Antibodies Inhibit Viral Egress and Protect from Lethal Influenza Infection in Mice
- PMID: 31757769
- PMCID: PMC6941661
- DOI: 10.1016/j.chom.2019.10.003
Influenza H7N9 Virus Neuraminidase-Specific Human Monoclonal Antibodies Inhibit Viral Egress and Protect from Lethal Influenza Infection in Mice
Abstract
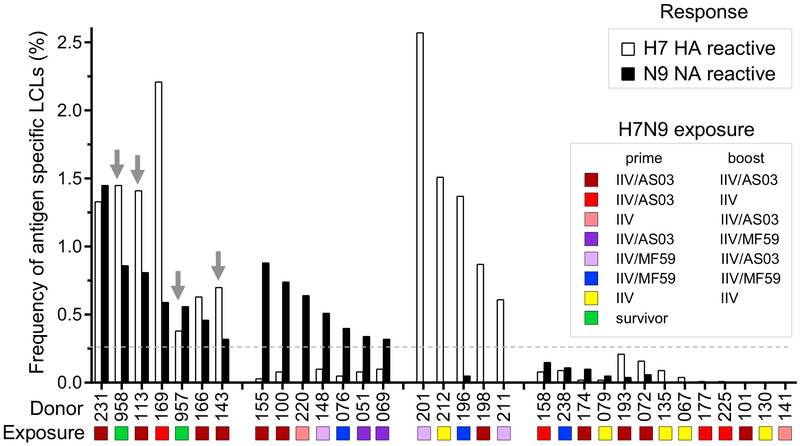
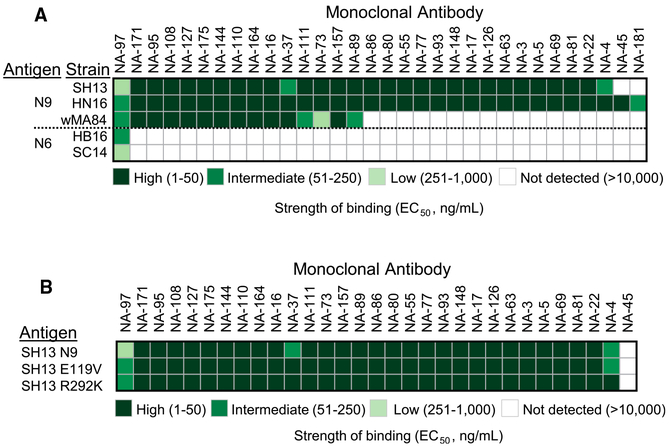
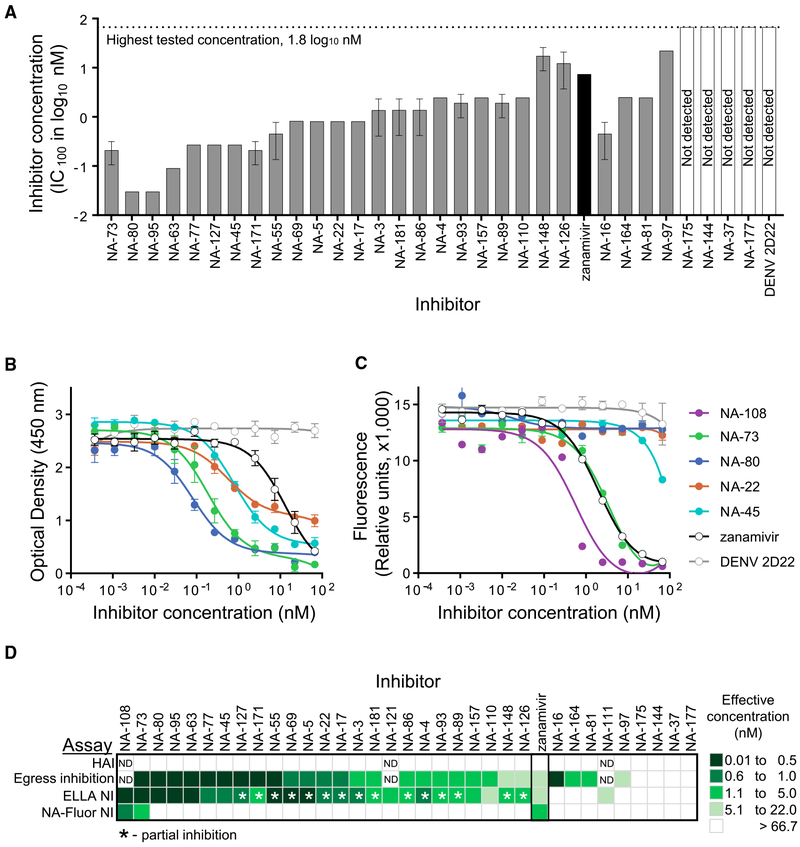
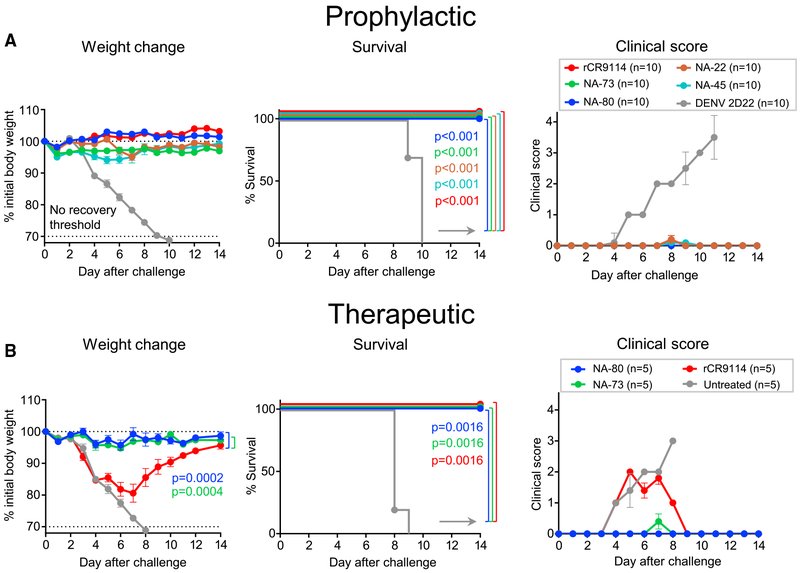
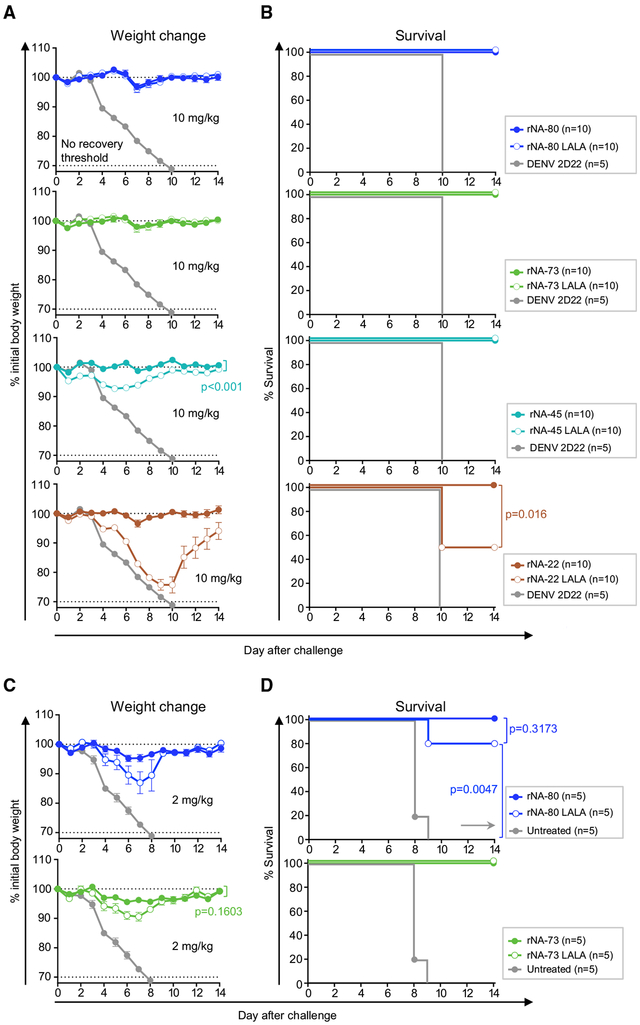
H7N9 avian influenza virus causes severe infections and might have the potential to trigger a major pandemic. Molecular determinants of human humoral immune response to N9 neuraminidase (NA) proteins, which exhibit unusual features compared with seasonal influenza virus NA proteins, are ill-defined. We isolated 35 human monoclonal antibodies (mAbs) from two H7N9 survivors and two vaccinees. These mAbs react to NA in a subtype-specific manner and recognize diverse antigenic sites on the surface of N9 NA, including epitopes overlapping with, or distinct from, the enzyme active site. Despite recognizing multiple antigenic sites, the mAbs use a common mechanism of action by blocking egress of nascent virions from infected cells, thereby providing an antiviral prophylactic and therapeutic protection in vivo in mice. Studies of breadth, potency, and diversity of antigenic recognition from four subjects suggest that vaccination with inactivated adjuvanted vaccine induce NA-reactive responses comparable to that of H7N9 natural infection.
Keywords: B lymphocyte; H7N9; antibodies; epitopes; influenza A virus; monoclonal; neuraminidase; pre-exposure prophylaxis.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
J.E.C. has served as a consultant for Takeda Vaccines, Sanofi Pasteur, Pfizer, and Novavax; is on the Scientific Advisory Boards of CompuVax and Meissa Vaccines; and is Founder of IDBiologics, Inc. All other authors declare no conflict of interest. Vanderbilt University has applied for a patent related to the NA antibodies.
Figures


Comment in
-
Emerging from the Shadow of Hemagglutinin: Neuraminidase Is an Important Target for Influenza Vaccination.Cell Host Microbe. 2019 Dec 11;26(6):712-713. doi: 10.1016/j.chom.2019.11.006. Cell Host Microbe. 2019. PMID: 31951584
References
-
- Arduin E, Arora S, Bamert PR, Kuiper T, Popp S, Geisse S, Grau R, Calzascia T, Zenke G, and Kovarik J (2015). Highly reduced binding to high and low affinity mouse Fc gamma receptors by L234A/L235A and N297A Fc mutations engineered into mouse IgG2a. Mol. Immunol 63, 456–463. - PubMed
-
- Beard KR, Brendish NJ, and Clark TW (2018).Treatment of influenza with neuraminidase inhibitors. Curr. Opin. Infect. Dis 31, 514–519. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
