

Transient Protein-RNA Interactions Guide Nascent Ribosomal RNA Folding
- PMID: 31761533
- PMCID: PMC7006226
- DOI: 10.1016/j.cell.2019.10.035
Transient Protein-RNA Interactions Guide Nascent Ribosomal RNA Folding
Abstract
Ribosome assembly is an efficient but complex and heterogeneous process during which ribosomal proteins assemble on the nascent rRNA during transcription. Understanding how the interplay between nascent RNA folding and protein binding determines the fate of transcripts remains a major challenge. Here, using single-molecule fluorescence microscopy, we follow assembly of the entire 3' domain of the bacterial small ribosomal subunit in real time. We find that co-transcriptional rRNA folding is complicated by the formation of long-range RNA interactions and that r-proteins self-chaperone the rRNA folding process prior to stable incorporation into a ribonucleoprotein (RNP) complex. Assembly is initiated by transient rather than stable protein binding, and the protein-RNA binding dynamics gradually decrease during assembly. This work questions the paradigm of strictly sequential and cooperative ribosome assembly and suggests that transient binding of RNA binding proteins to cellular RNAs could provide a general mechanism to shape nascent RNA folding during RNP assembly.
Keywords: RNA biology; RNA chaperones; RNA misfolding; co-transcriptional RNA folding; cooperativity; protein-RNA dynamics; protein-RNA interactions; ribonucleoprotein assembly; ribosome assembly; single-molecule fluorescence microscopy.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
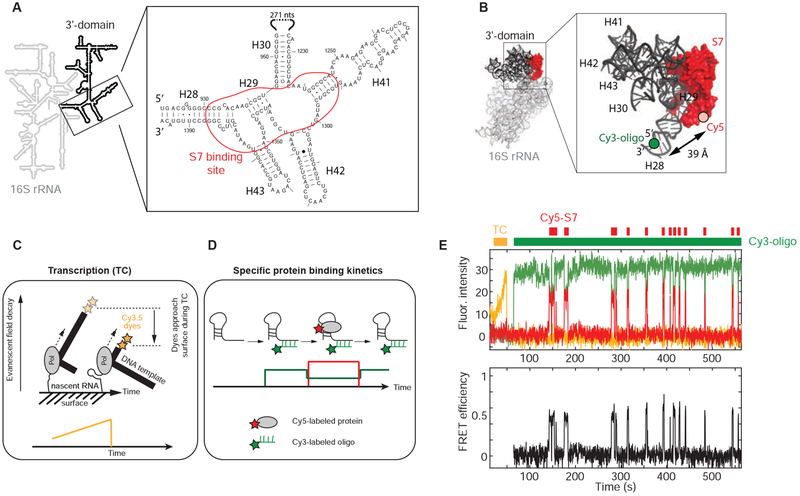
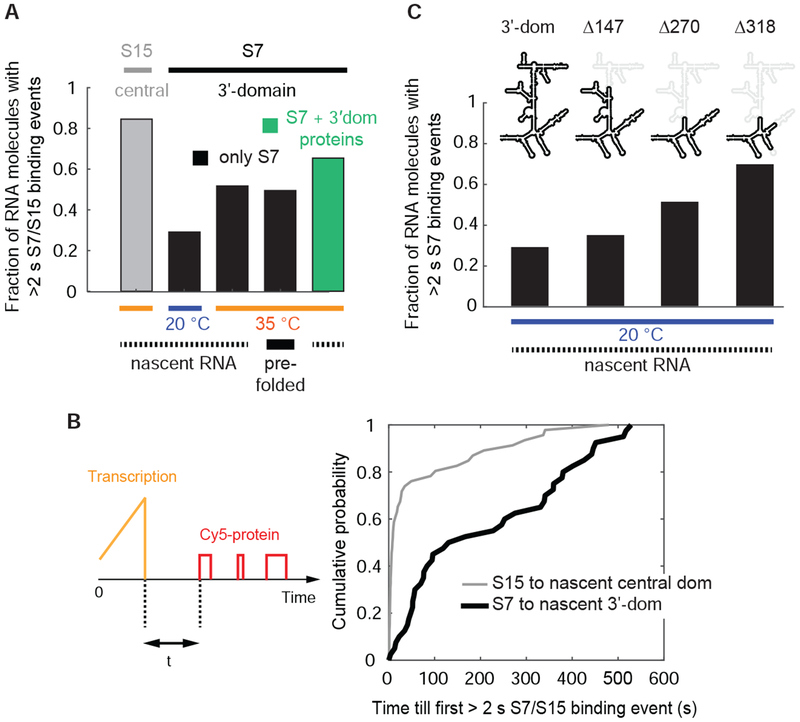
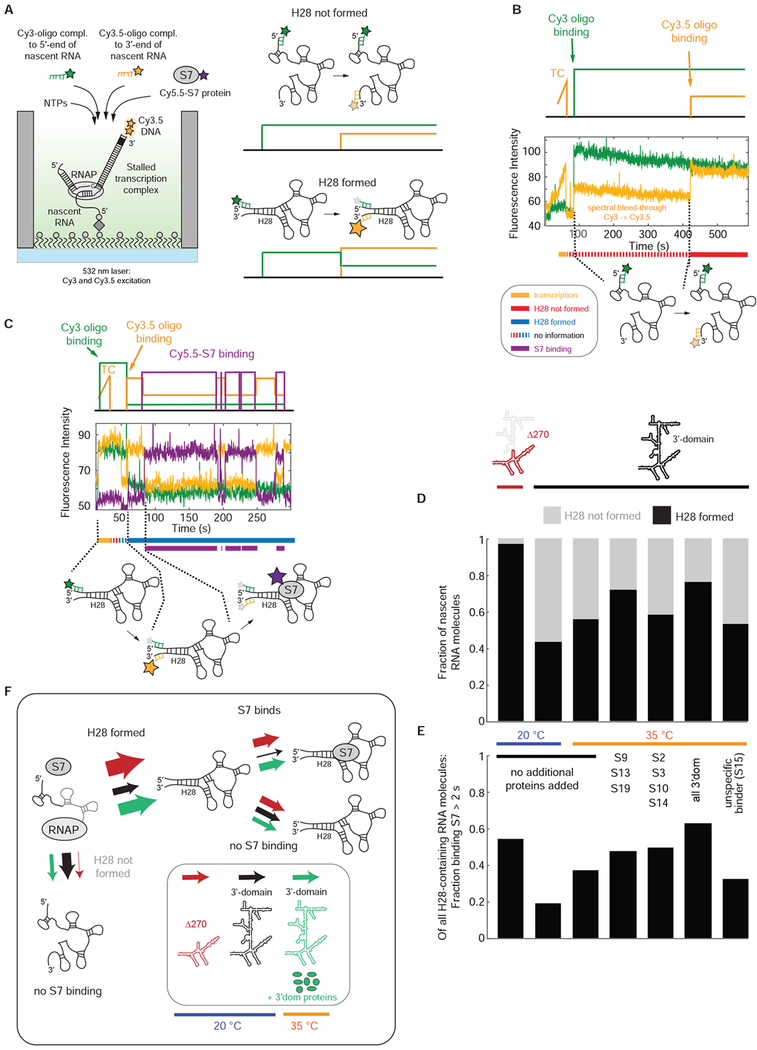
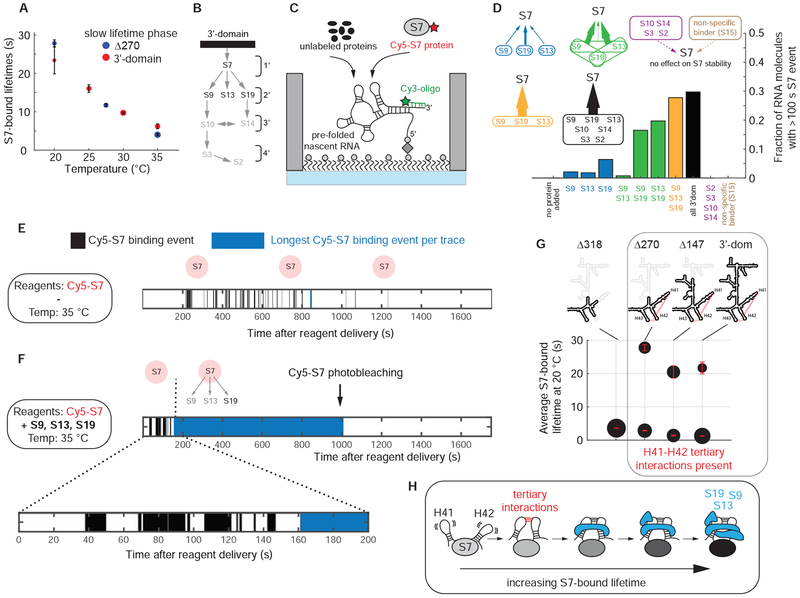
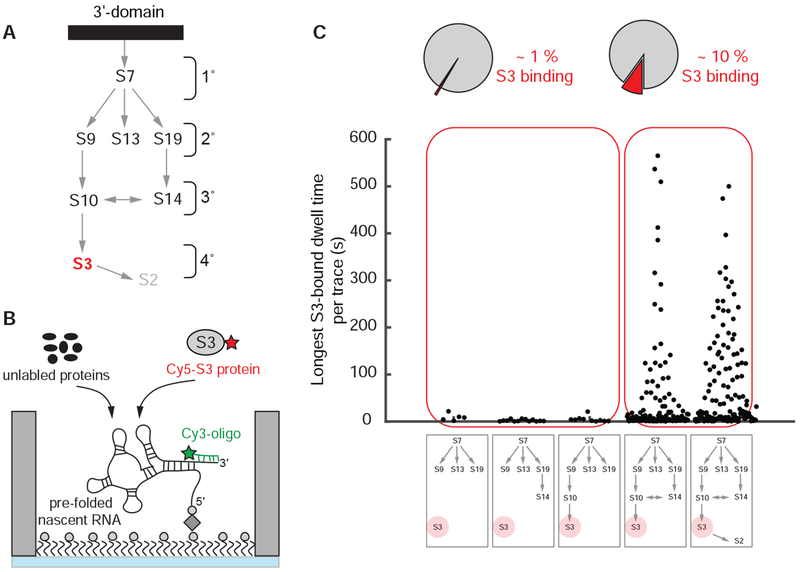
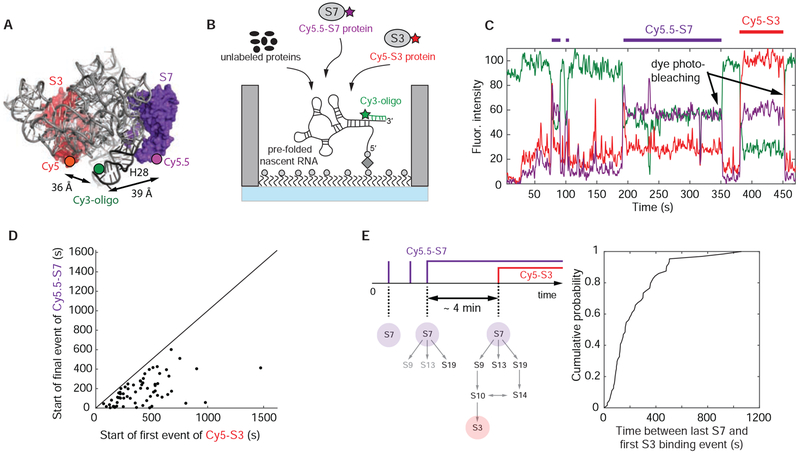
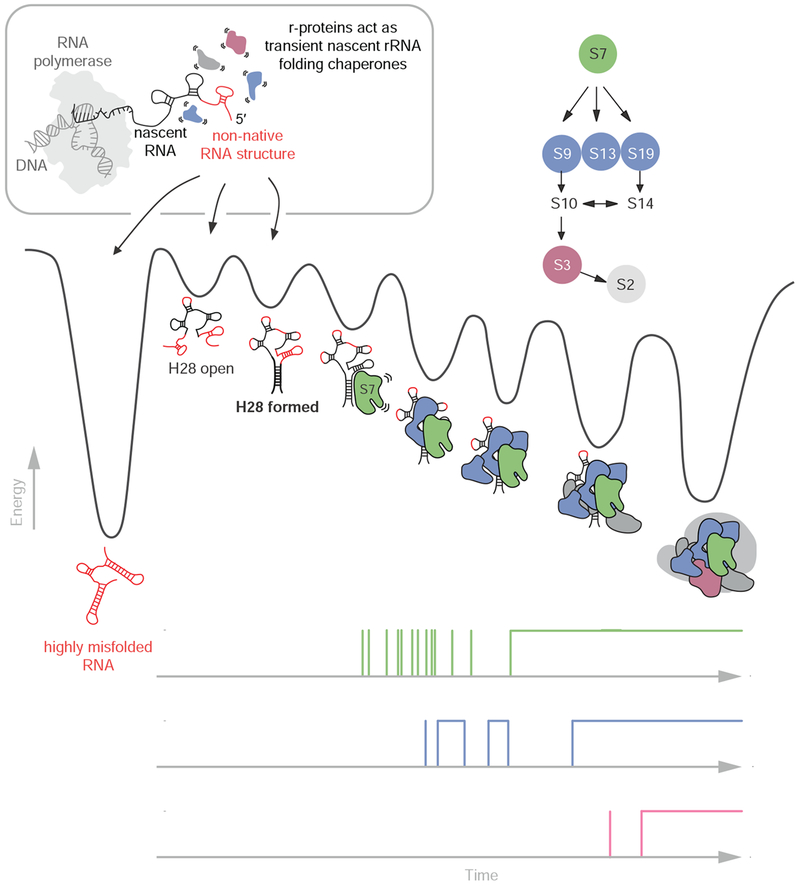
Comment in
-
RNA-Binding Proteins Chaperone Ribonucleoprotein Complex Assembly to Solve the RNA-Folding Problem.Cell. 2019 Nov 27;179(6):1248-1250. doi: 10.1016/j.cell.2019.11.011. Epub 2019 Nov 21. Cell. 2019. PMID: 31761531
References
-
- Boyle J, Robillard GT, and Kim SH (1980). Sequential folding of transfer RNA. A nuclear magnetic resonance study of successively longer tRNA fragments with a common 5′ end. J Mol Biol 139, 601–625. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
