

Characterizing how probiotic Lactobacillus reuteri 6475 and lactobacillic acid mediate suppression of osteoclast differentiation
- PMID: 31763377
- PMCID: PMC6864341
- DOI: 10.1016/j.bonr.2019.100227
Characterizing how probiotic Lactobacillus reuteri 6475 and lactobacillic acid mediate suppression of osteoclast differentiation
Abstract
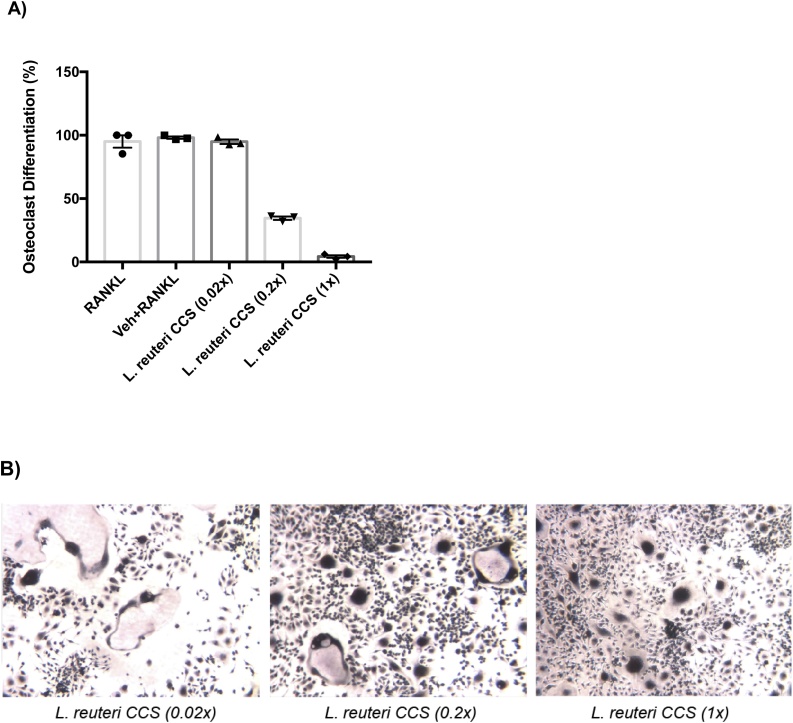
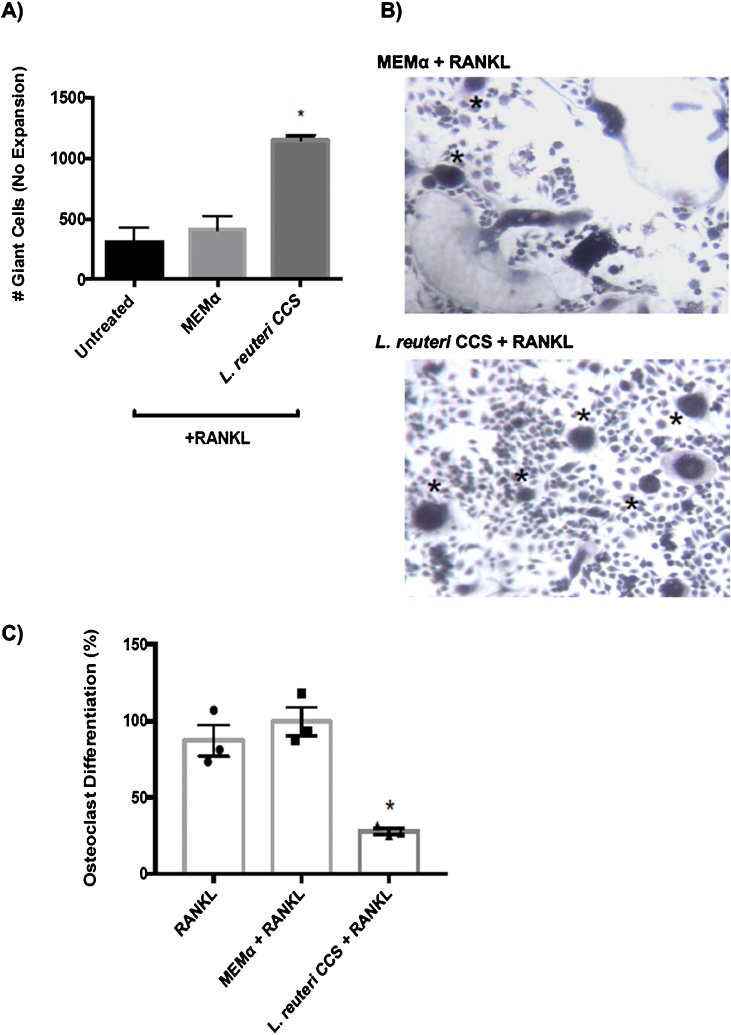
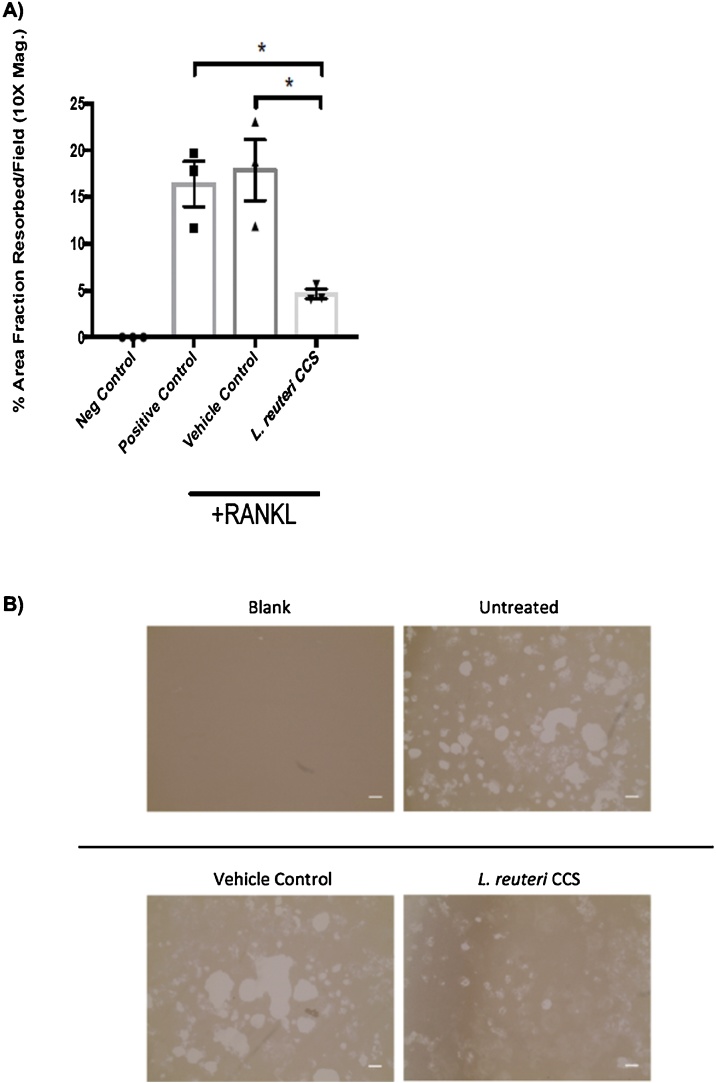
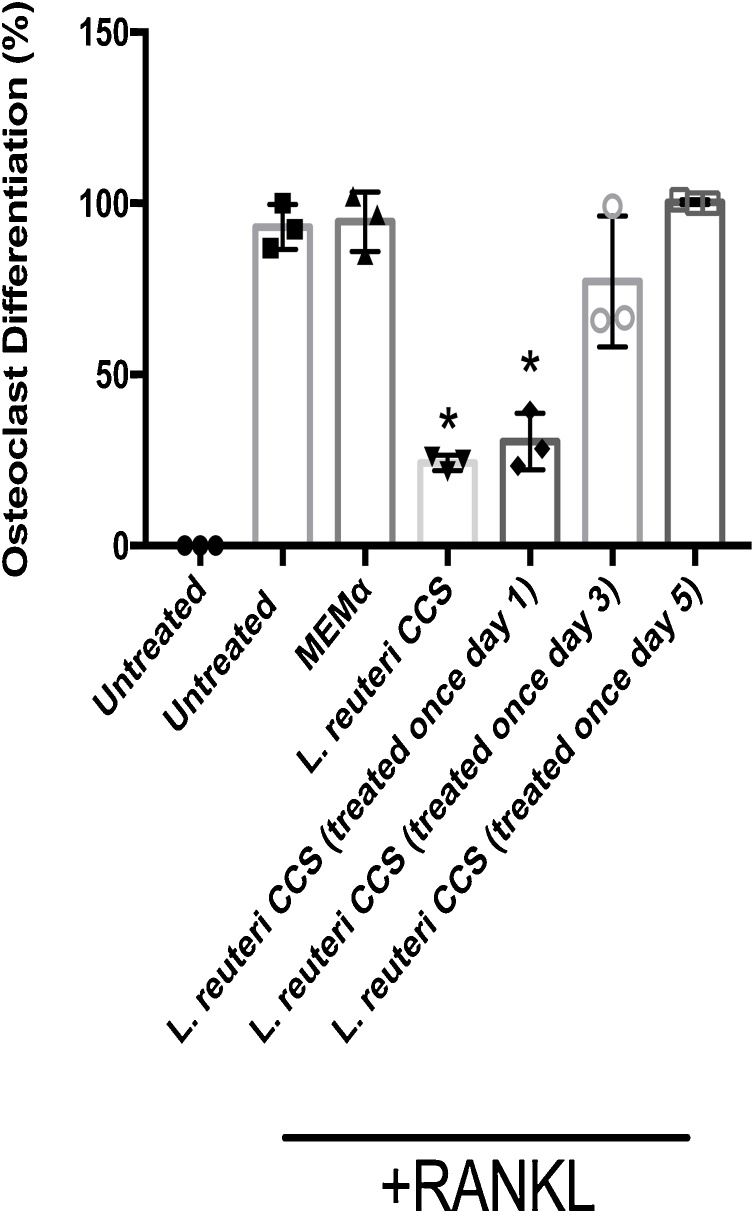
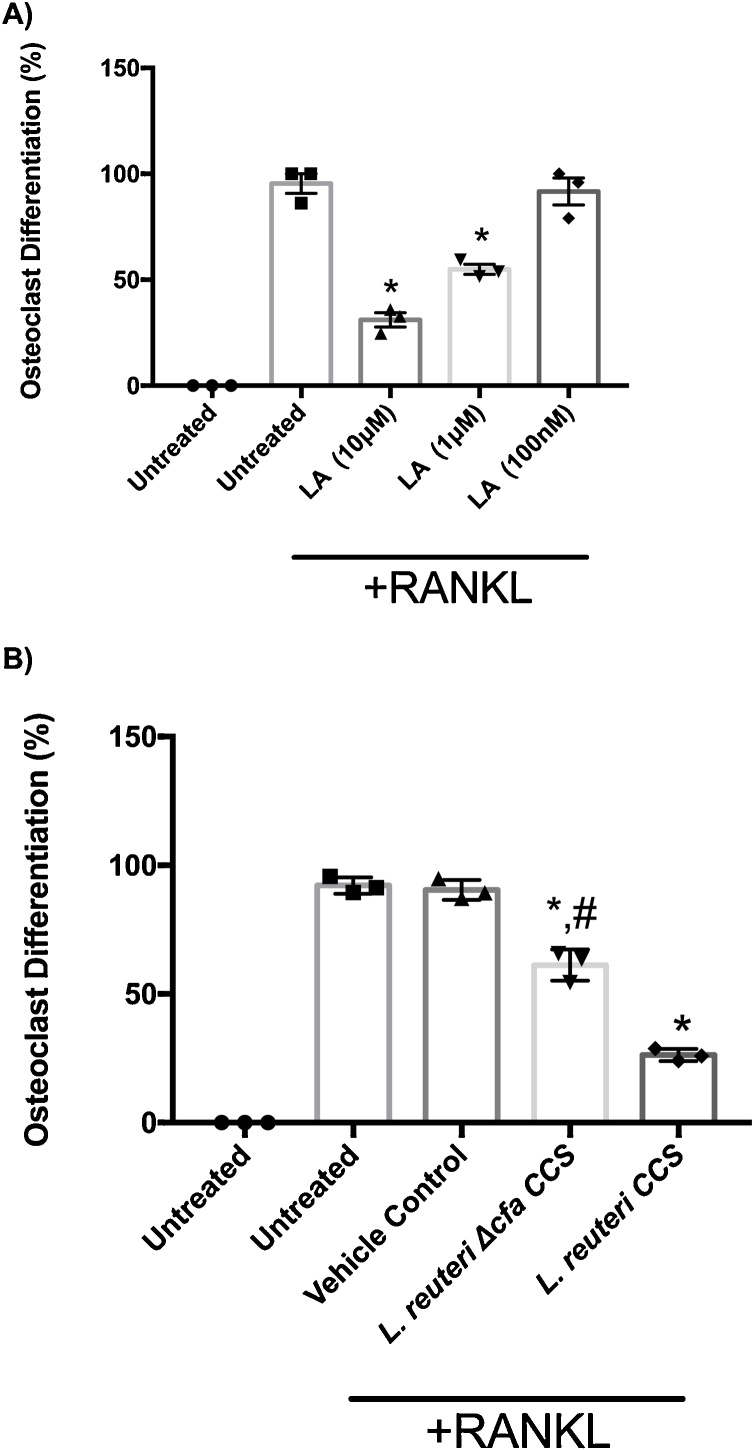
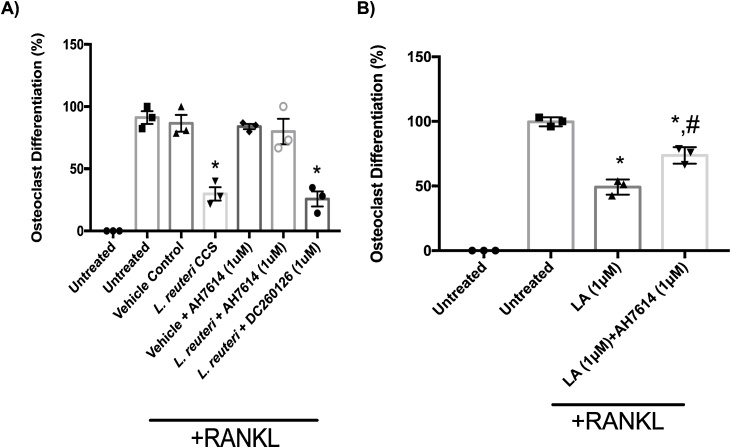
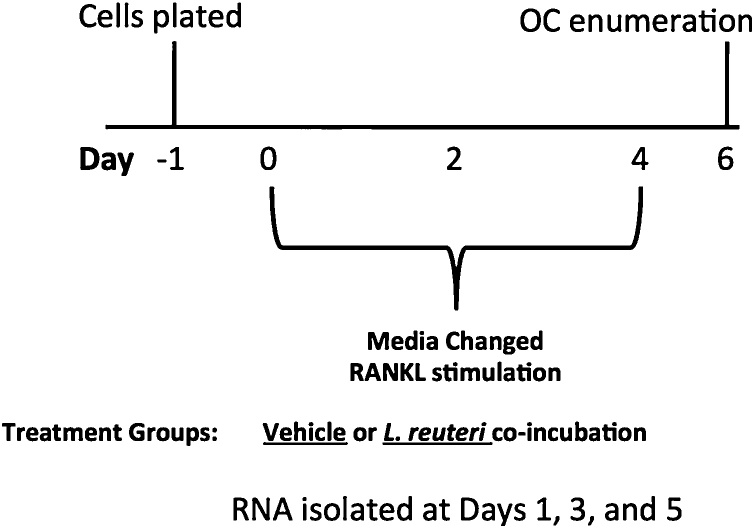
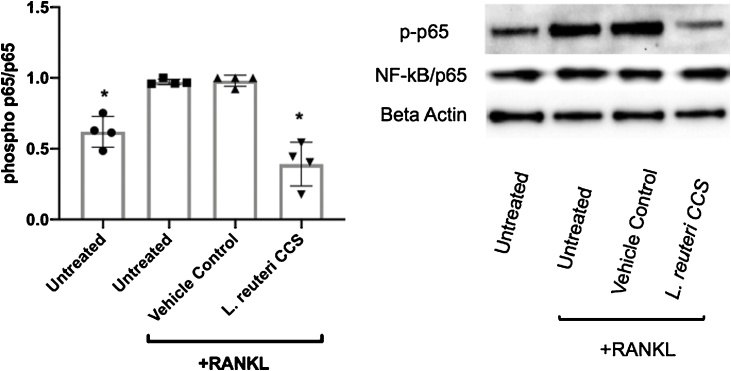
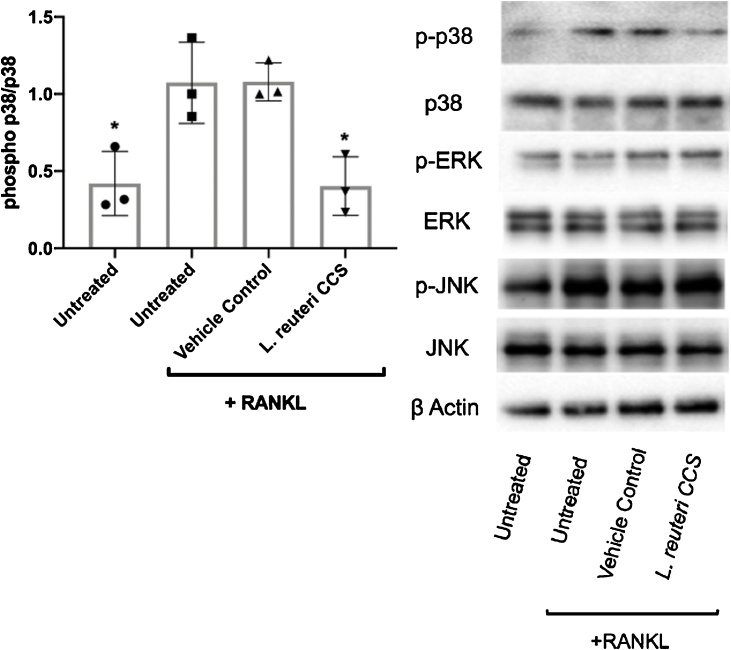
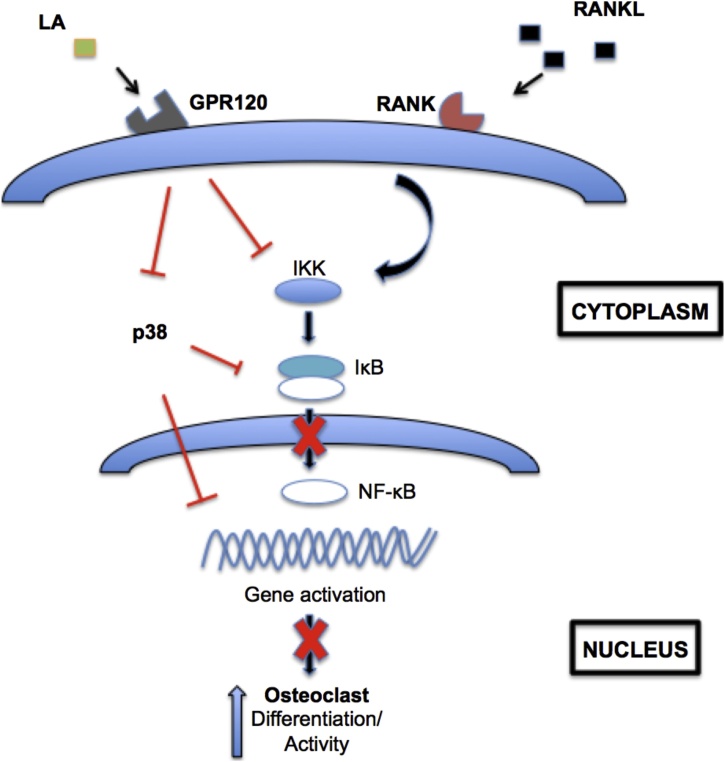
Osteoporosis is a disease that impacts over 200 million people worldwide. Taking into consideration the side effects stemming from medications used to treat this illness, investigators have increased their efforts to develop novel therapeutics for osteoporosis. In a previous study, we demonstrated that ovariectomy-induced bone loss in mice was prevented by treatment with the probiotic bacterium Lactobacillus reuteri 6475 (L. reuteri), an effect that correlated with reduced osteoclastogenesis in the bone marrow of L. reuteri treated mice. We also demonstrated that L. reuteri directly inhibited osteoclastogenesis in vitro. To better understand how L. reuteri impacts osteoclast formation, we used additional in vitro analyses to identify that conditioned supernatant from L. reuteri inhibited osteoclastogenesis at the intermediate stage of fused polykaryons. To elucidate the effect of L. reuteri treatment on host cell physiology, we performed RNAseq at multiple time points during in vitro osteoclastogenesis and established that L. reuteri downregulated several KEGG pathways including osteoclast differentiation as well as TNF-α, NF-κB, and MAP kinase signaling. These results were consistent with Western Blot data demonstrating that NF-κB and p38 activation were decreased by L. reuteri treatment. We further identified that lactobacillic acid (LA), a cyclopropane fatty acid produced by L. reuteri, contributed significantly to the suppression of osteoclastogenesis. Additionally, we demonstrated that L. reuteri is signaling through the long chain fatty acid receptor, GPR120, to impact osteoclastogenesis. Overall, these studies provide both bacterial and host mechanisms by which L. reuteri impacts osteoclastogenesis and suggest that long chain fatty acid receptors could be targets for preventing osteoclastogenesis.
Keywords: Bone; Lactobacillic acid; Osteoclast; Osteoporosis; Probiotic.
© 2019 The Author(s).
Conflict of interest statement
The authors report that they have no competing interests.
Figures
References
-
- Aharon R., Bar-Shavit Z. Involvement of aquaporin 9 in osteoclast differentiation. J. Biol. Chem. 2006;281:19305–19309. - PubMed
-
- Ahn S.H., Park S.-Y., Baek J.-E., Lee S.-Y., Baek W.-Y., Lee S.-Y., Lee Y.-S., Ju Yoo H., Kim H., Lee S.H., Im D.-S., Lee S.-K., Kim B.-J., Koh J.-M. Free fatty acid receptor 4 (GPR120) stimulates bone formation and suppresses bone resorption in the presence of elevated n -3 fatty acid levels. Endocrinology. 2016;4 - PubMed
-
- Anders S., McCarthy D.J., Chen Y., Okoniewski M., Smyth G.K., Huber W., Robinson M.D. Count-based differential expression analysis of RNA sequencing data using R and Bioconductor. Nat. Protoc. 2013;8:1765–1786. doi:nprot.2013.099[pii]\r10.1038/nprot.2013.099. - PubMed
-
- Arthur J.S., Ley S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013;13:679–692. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
