

Insulin-like growth factor (IGF)-II- mediated fibrosis in pathogenic lung conditions
- PMID: 31765403
- PMCID: PMC6876936
- DOI: 10.1371/journal.pone.0225422
Insulin-like growth factor (IGF)-II- mediated fibrosis in pathogenic lung conditions
Abstract
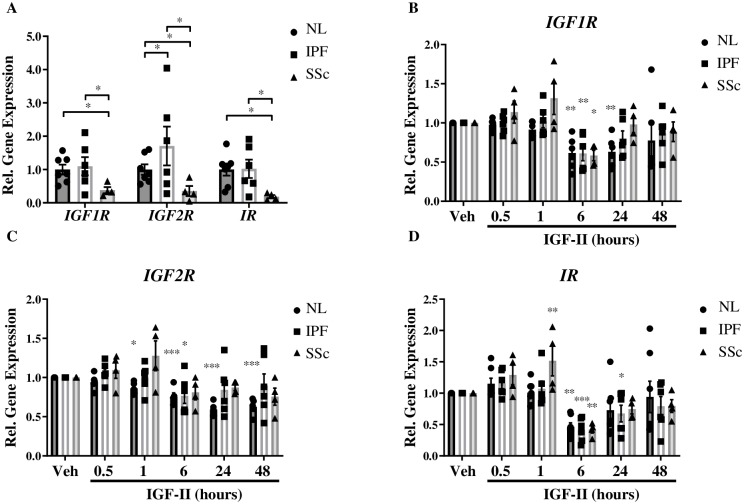
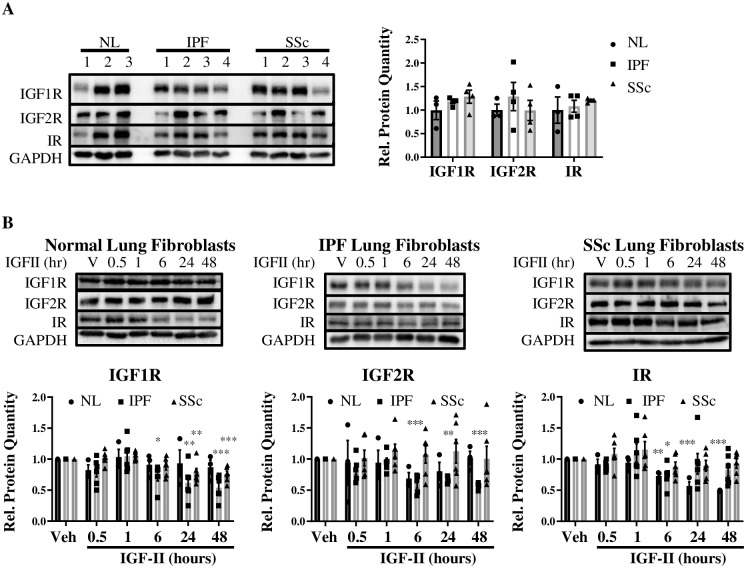
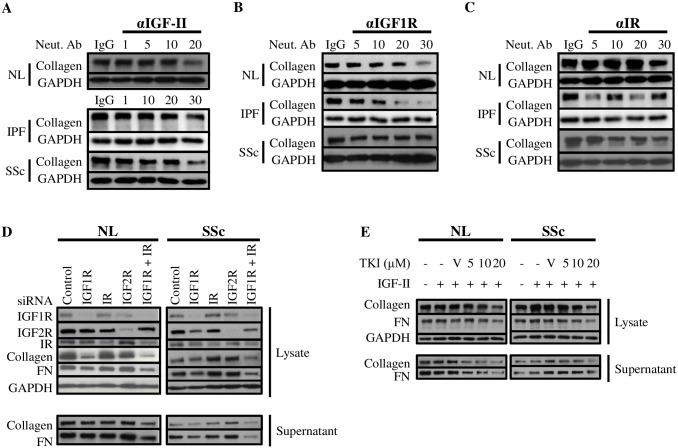
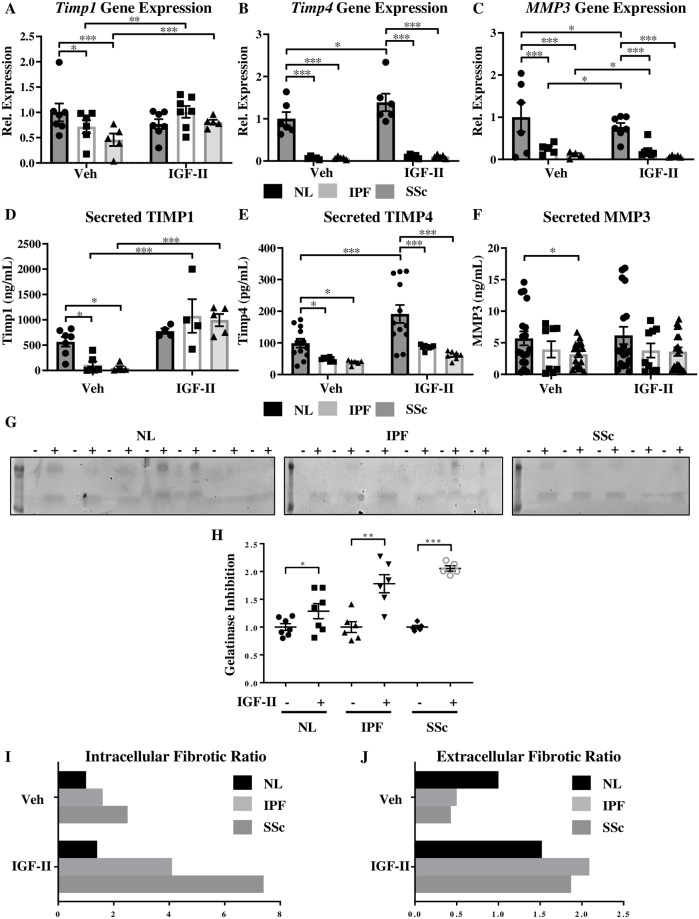
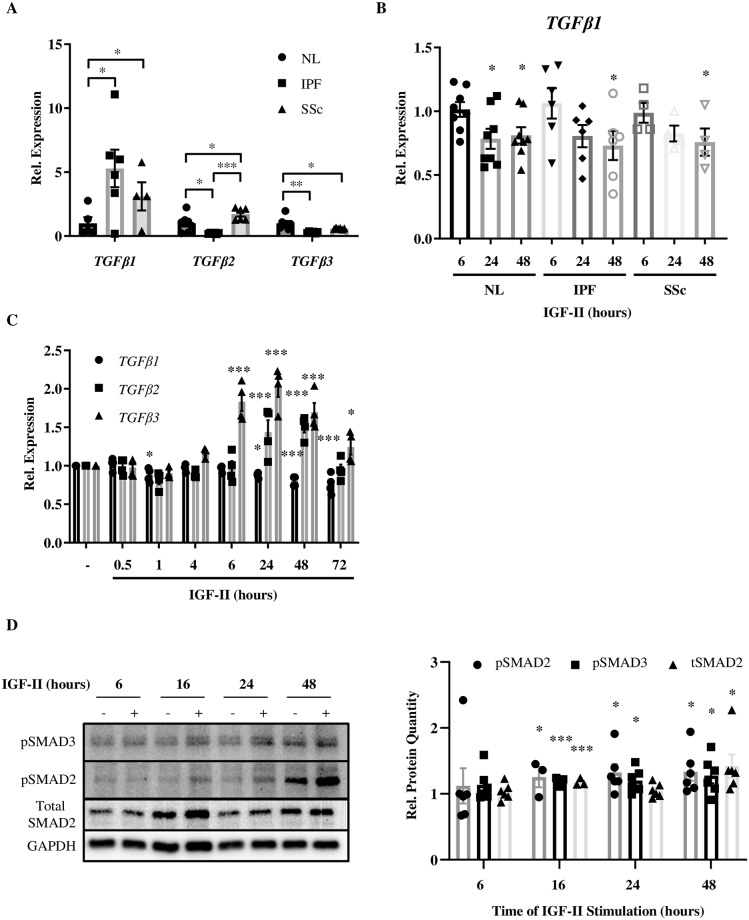
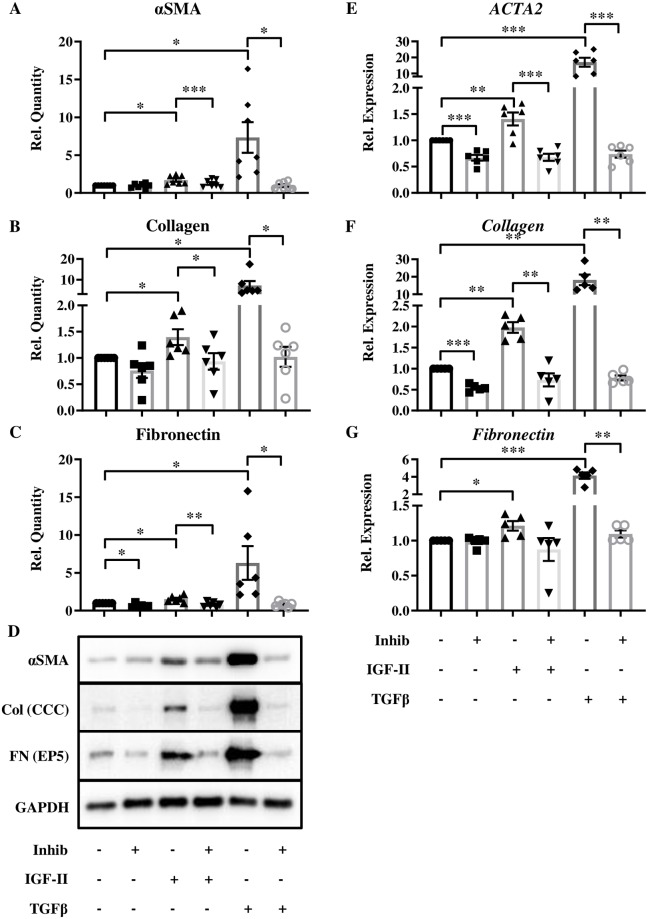
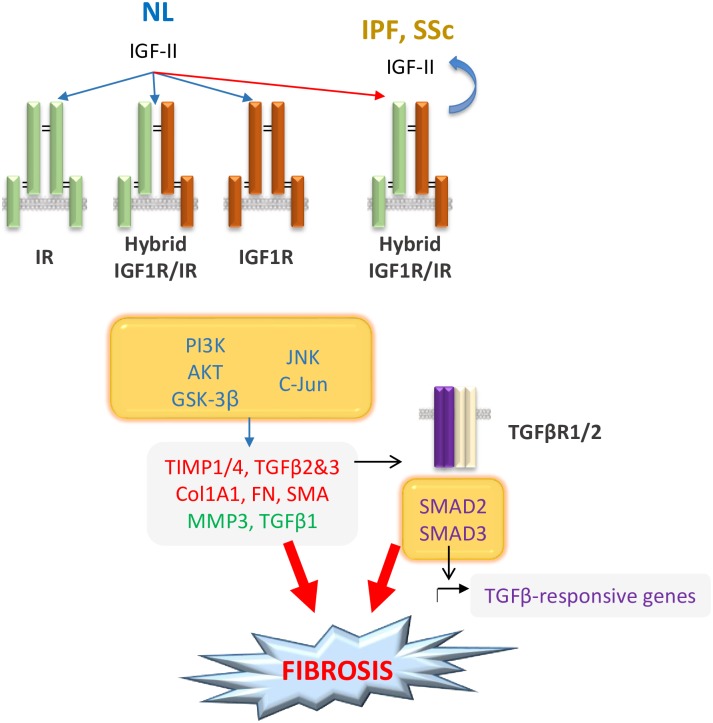
Type 2 insulin-like growth factor (IGF-II) levels are increased in fibrosing lung diseases such as idiopathic pulmonary fibrosis (IPF) and scleroderma/systemic sclerosis-associated pulmonary fibrosis (SSc). Our goal was to investigate the contribution of IGF receptors to IGF-II-mediated fibrosis in these diseases and identify other potential mechanisms key to the fibrotic process. Cognate receptor gene and protein expression were analyzed with qRT-PCR and immunoblot in primary fibroblasts derived from lung tissues of normal donors (NL) and patients with IPF or SSc. Compared to NL, steady-state receptor gene expression was decreased in SSc but not in IPF. IGF-II stimulation differentially decreased receptor mRNA and protein levels in NL, IPF, and SSc fibroblasts. Neutralizing antibody, siRNA, and receptor inhibition targeting endogenous IGF-II and its primary receptors, type 1 IGF receptor (IGF1R), IGF2R, and insulin receptor (IR) resulted in loss of the IGF-II response. IGF-II tipped the TIMP:MMP balance, promoting a fibrotic environment both intracellularly and extracellularly. Differentiation of fibroblasts into myofibroblasts by IGF-II was blocked with a TGFβ1 receptor inhibitor. IGF-II also increased TGFβ2 and TGFβ3 expression, with subsequent activation of canonical SMAD2/3 signaling. Therefore, IGF-II promoted fibrosis through IGF1R, IR, and IGF1R/IR, differentiated fibroblasts into myofibroblasts, decreased protease production and extracellular matrix degradation, and stimulated expression of two TGFβ isoforms, suggesting that IGF-II exerts pro-fibrotic effects via multiple mechanisms.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
