

Structure-Function Analysis Reveals the Singularity of Plant Mitochondrial DNA Replication Components: A Mosaic and Redundant System
- PMID: 31766564
- PMCID: PMC6963530
- DOI: 10.3390/plants8120533
Structure-Function Analysis Reveals the Singularity of Plant Mitochondrial DNA Replication Components: A Mosaic and Redundant System
Abstract
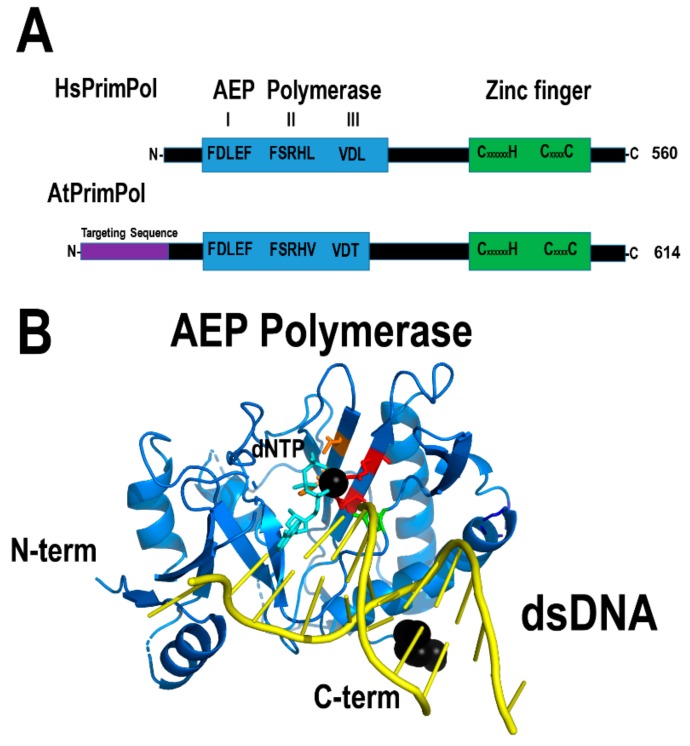
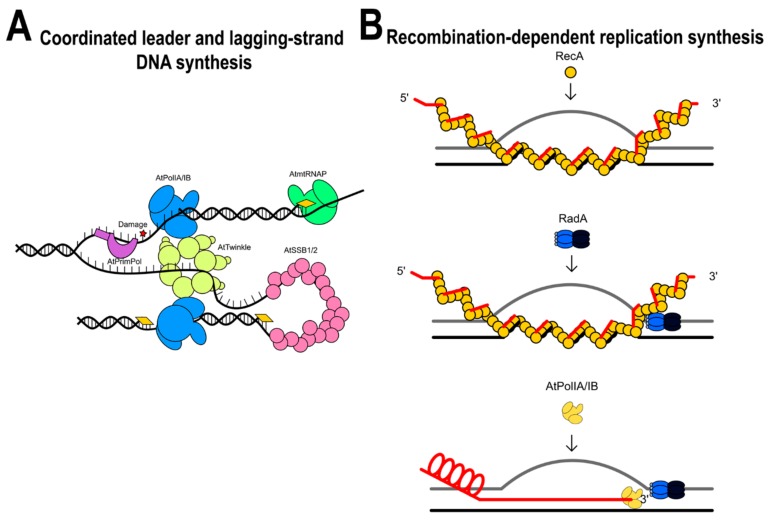
Plants are sessile organisms, and their DNA is particularly exposed to damaging agents. The integrity of plant mitochondrial and plastid genomes is necessary for cell survival. During evolution, plants have evolved mechanisms to replicate their mitochondrial genomes while minimizing the effects of DNA damaging agents. The recombinogenic character of plant mitochondrial DNA, absence of defined origins of replication, and its linear structure suggest that mitochondrial DNA replication is achieved by a recombination-dependent replication mechanism. Here, I review the mitochondrial proteins possibly involved in mitochondrial DNA replication from a structural point of view. A revision of these proteins supports the idea that mitochondrial DNA replication could be replicated by several processes. The analysis indicates that DNA replication in plant mitochondria could be achieved by a recombination-dependent replication mechanism, but also by a replisome in which primers are synthesized by three different enzymes: Mitochondrial RNA polymerase, Primase-Helicase, and Primase-Polymerase. The recombination-dependent replication model and primers synthesized by the Primase-Polymerase may be responsible for the presence of genomic rearrangements in plant mitochondria.
Keywords: DNA replication 1; evolution 2; recombination-dependent replication 4; replisome 3.
Conflict of interest statement
The author declares no conflict of interest.
Figures
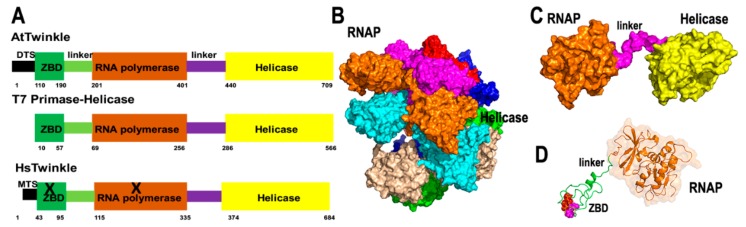
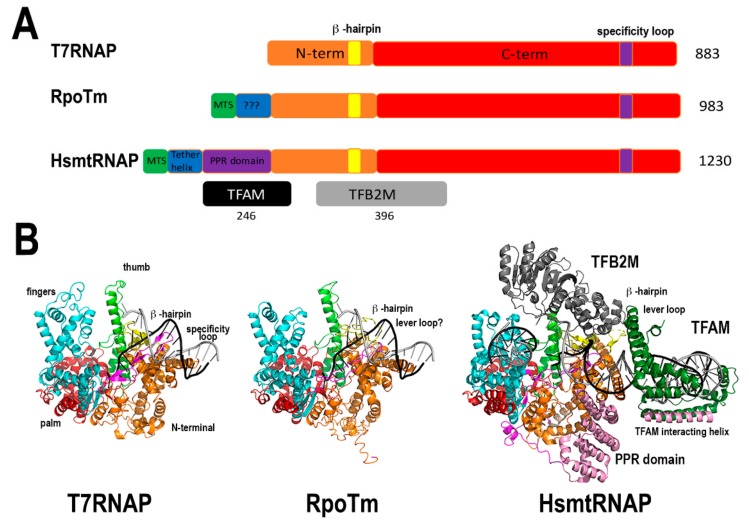
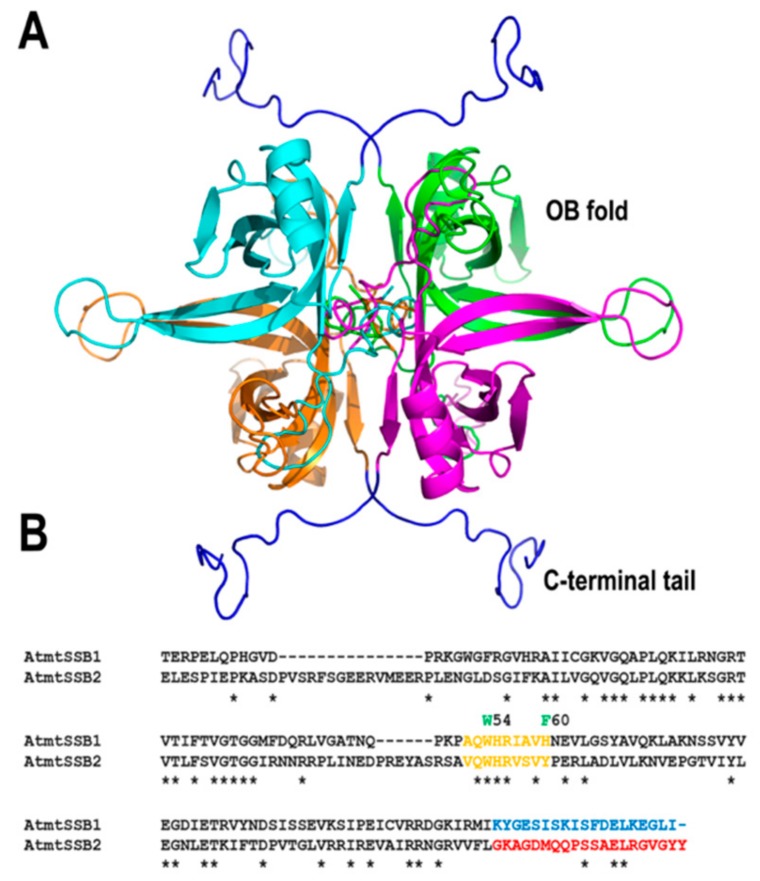
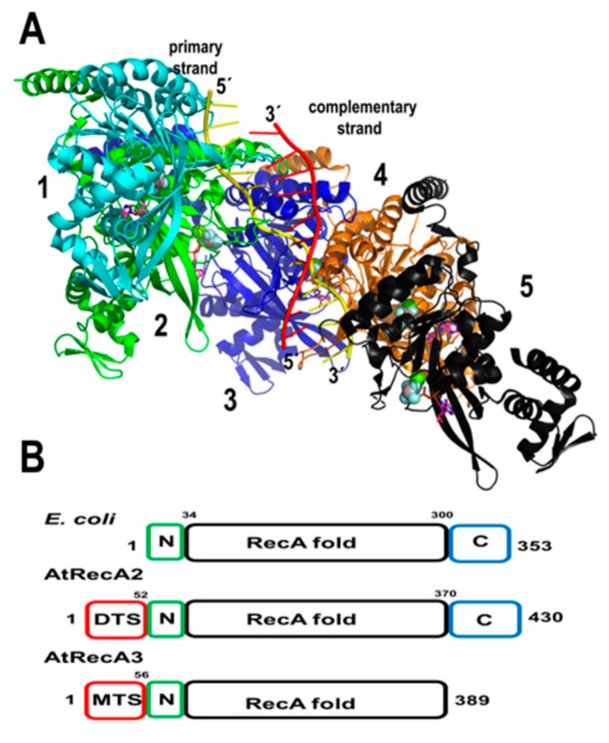
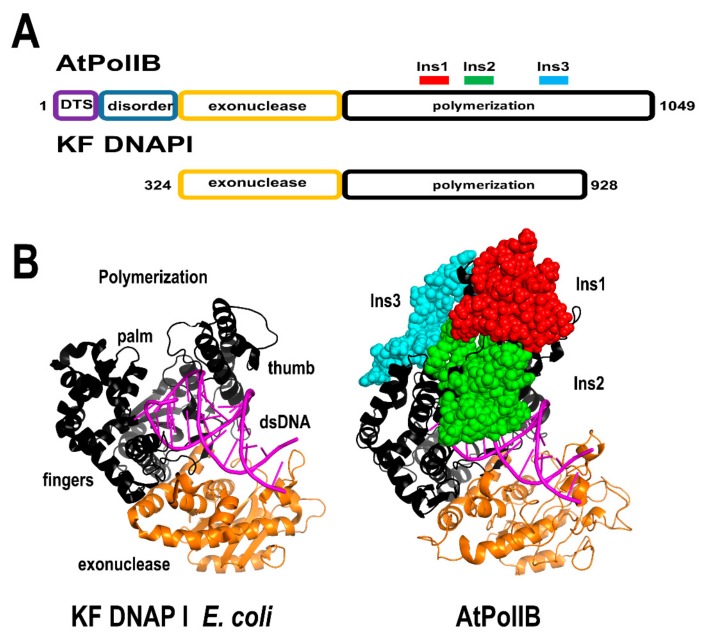
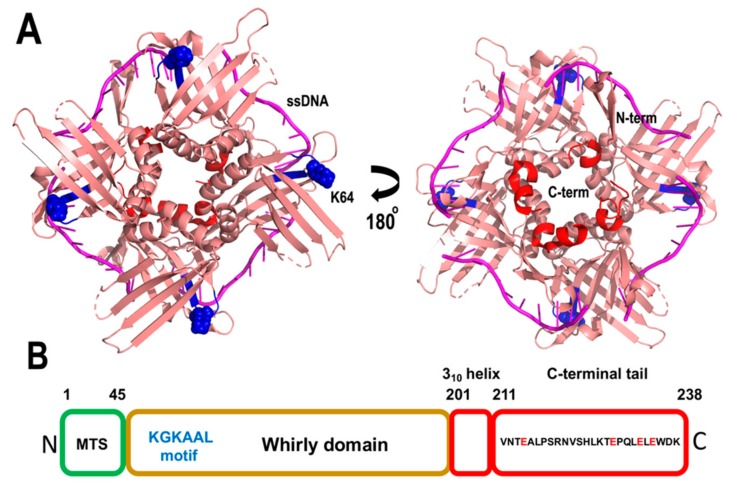
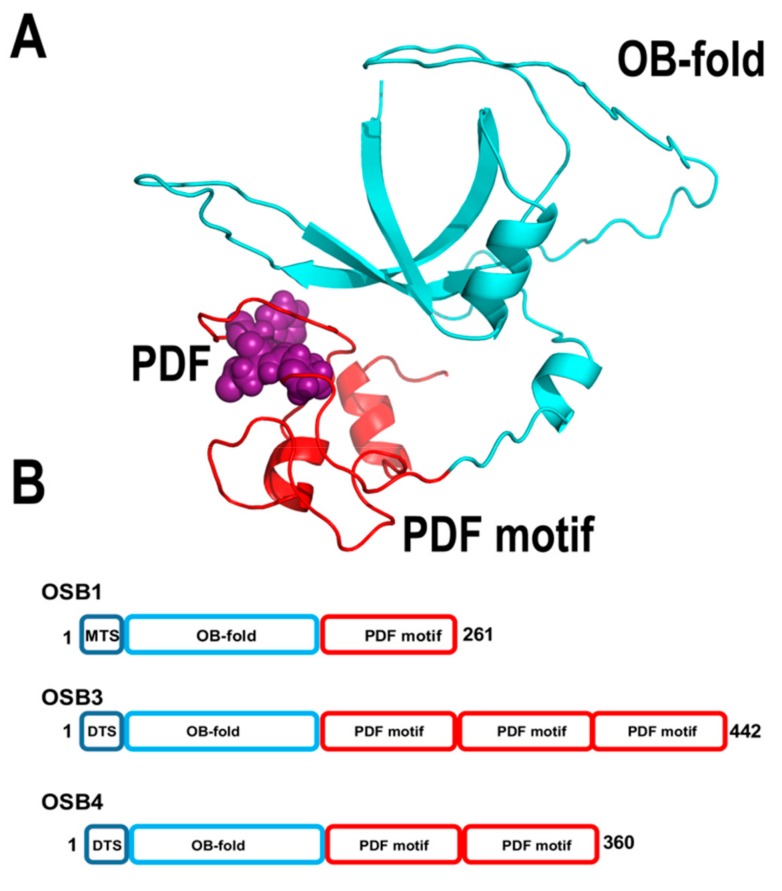
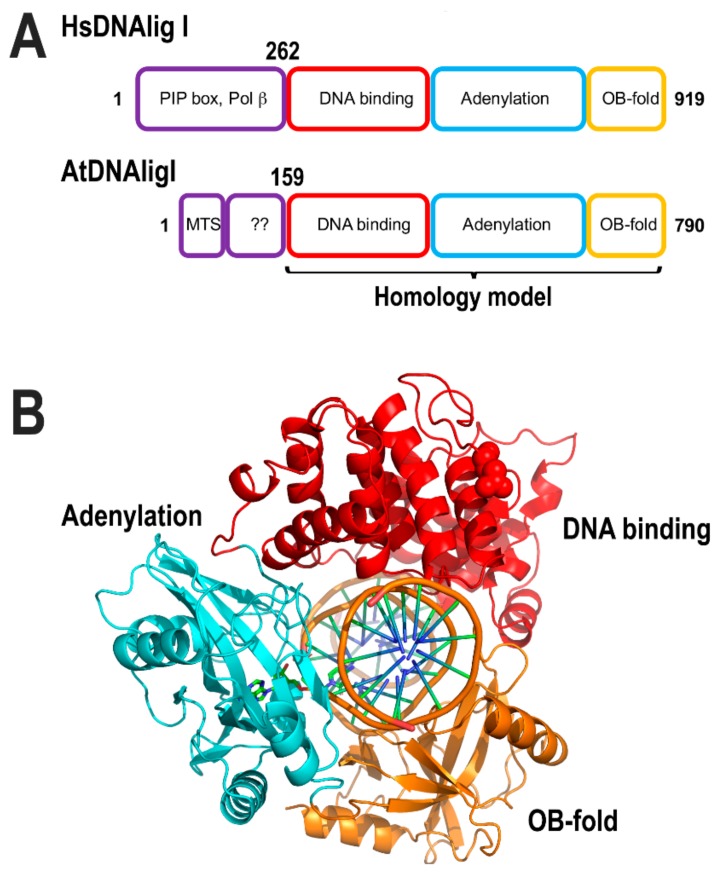
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
