

Regulation of Mitochondrial ATP Production: Ca2+ Signaling and Quality Control
- PMID: 31767352
- PMCID: PMC7921598
- DOI: 10.1016/j.molmed.2019.10.007
Regulation of Mitochondrial ATP Production: Ca2+ Signaling and Quality Control
Abstract
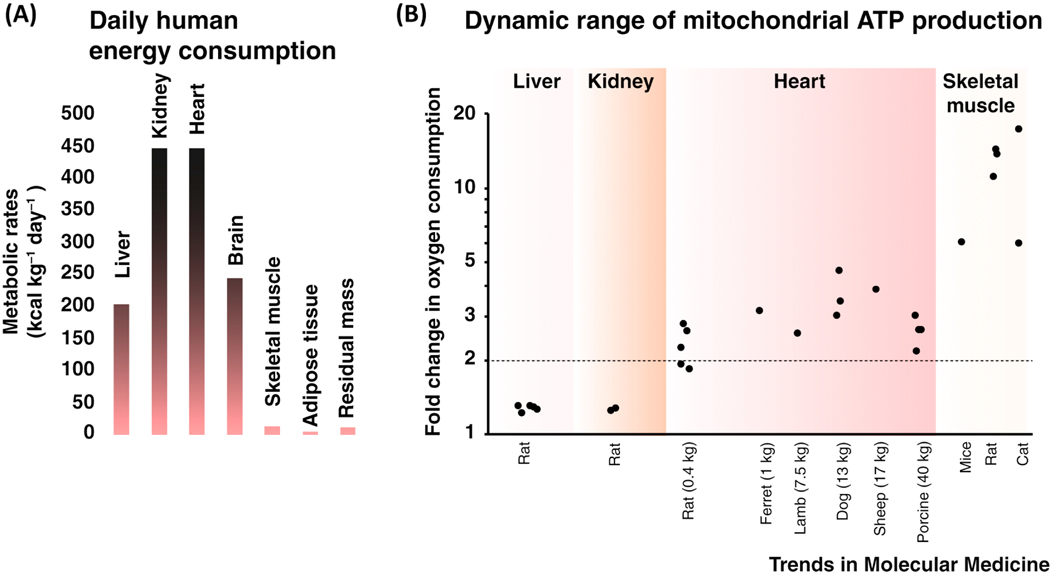
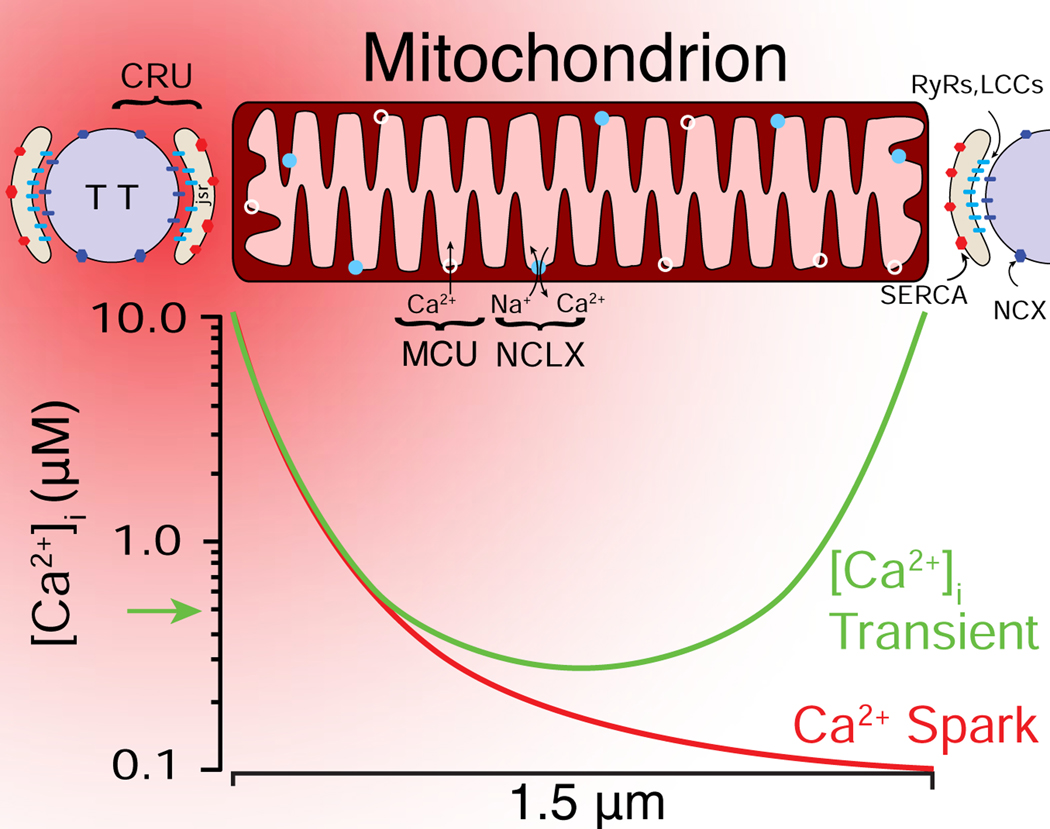
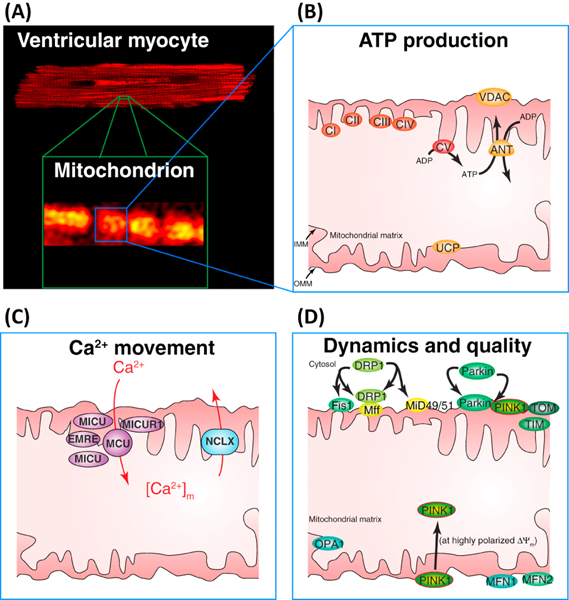
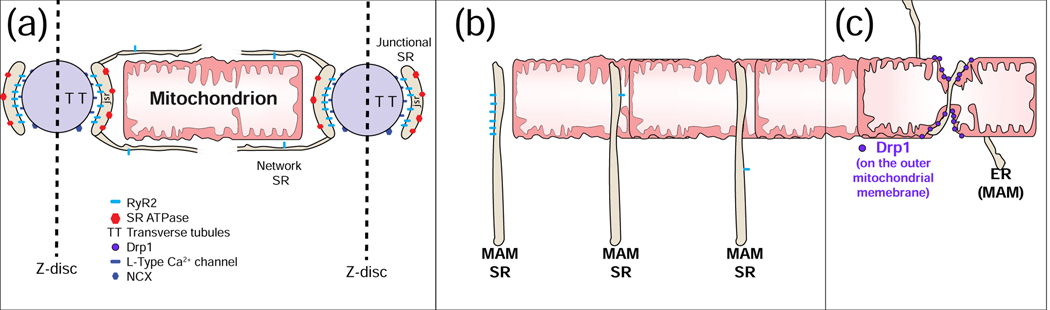
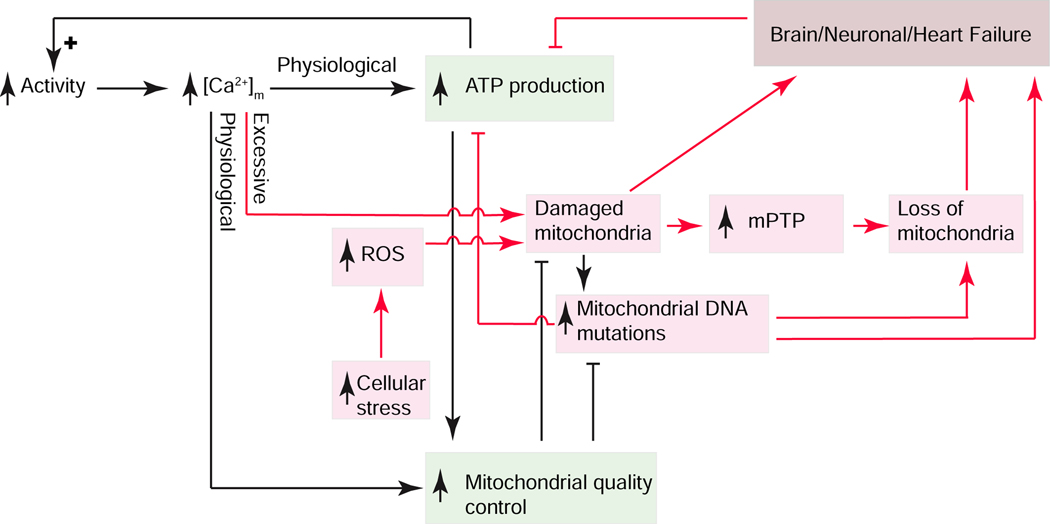
Cardiac ATP production primarily depends on oxidative phosphorylation in mitochondria and is dynamically regulated by Ca2+ levels in the mitochondrial matrix as well as by cytosolic ADP. We discuss mitochondrial Ca2+ signaling and its dysfunction which has recently been linked to cardiac pathologies including arrhythmia and heart failure. Similar dysfunction in other excitable and long-lived cells including neurons is associated with neurodegenerative diseases such as Alzheimer's disease (AD), amyotrophic lateral sclerosis (ALS), and Parkinson's disease (PD). Central to this new understanding is crucial Ca2+ regulation of both mitochondrial quality control and ATP production. Mitochondria-associated membrane (MAM) signaling from the sarcoplasmic reticulum (SR) and the endoplasmic reticulum (ER) to mitochondria is discussed. We propose future research directions that emphasize a need to define quantitatively the physiological roles of MAMs, as well as mitochondrial quality control and ATP production.
Keywords: heart failure; mitochondria-associated membranes; mitochondrial Ca(2+) signaling; mitochondrial quality control; mitophagy; neurodegeneration.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures
References
-
- McCormack JG and Denton RM (1993) The role of intramitochondrial Ca2+ in the regulation of oxidative phosphorylation in mammalian tissues. Biochem Soc Trans 21 ( Pt 3) (3), 793–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
