

Nucleolar dynamics and interactions with nucleoplasm in living cells
- PMID: 31769409
- PMCID: PMC6879204
- DOI: 10.7554/eLife.47533
Nucleolar dynamics and interactions with nucleoplasm in living cells
Abstract
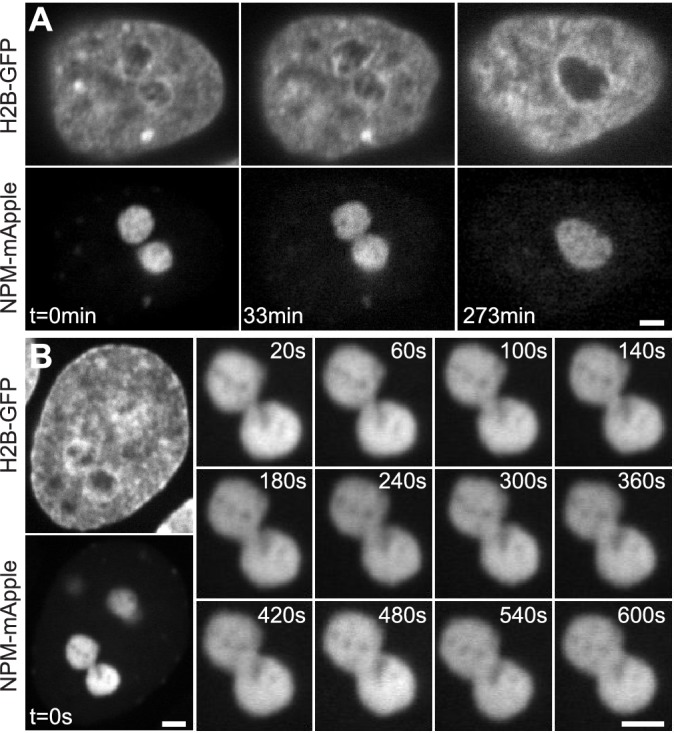
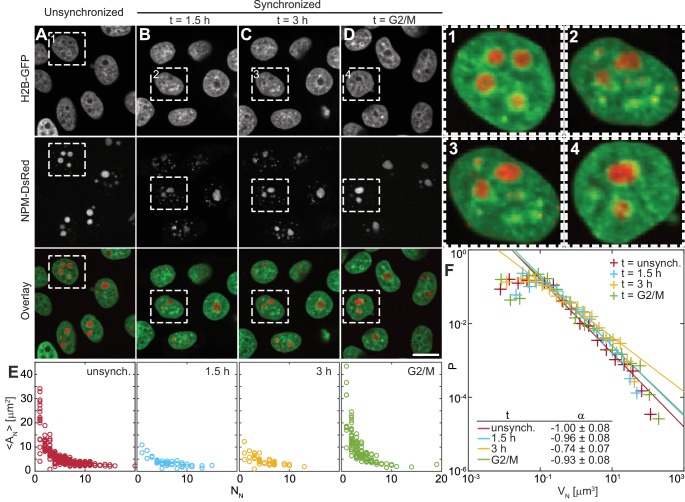
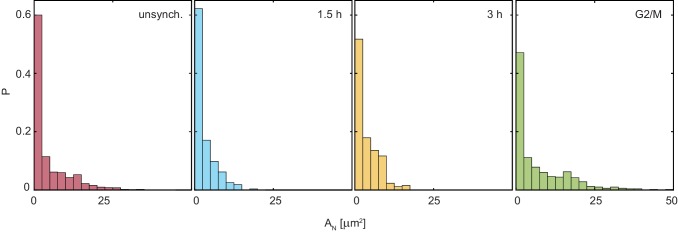
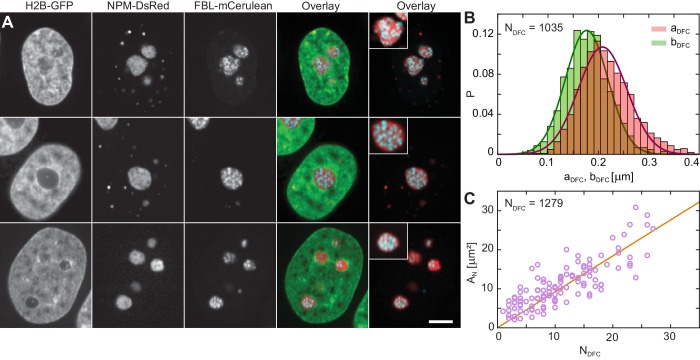
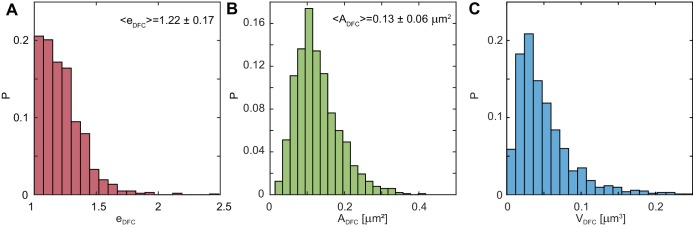
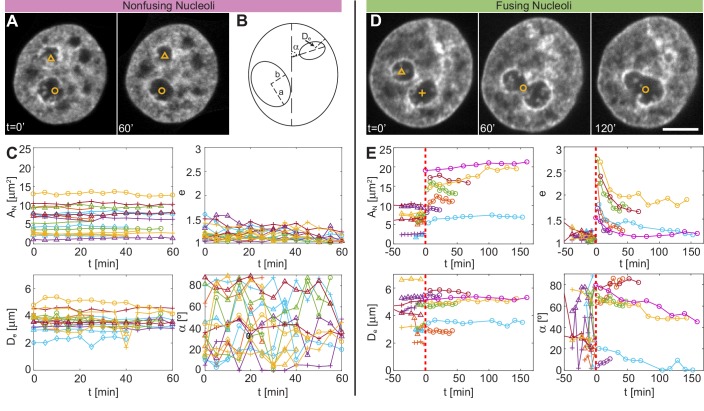
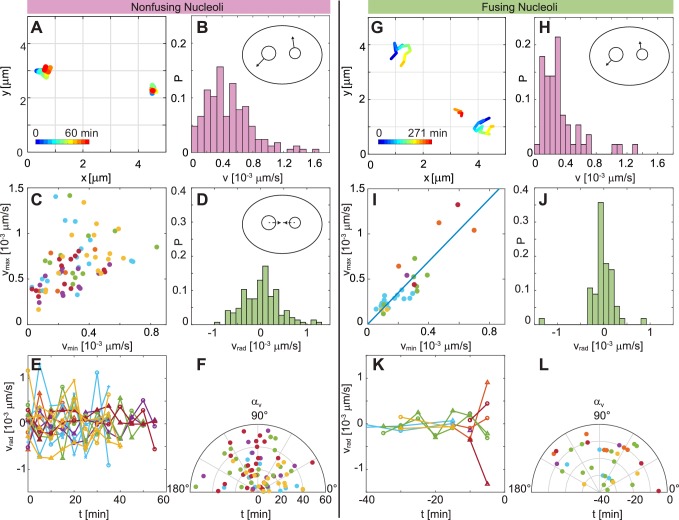
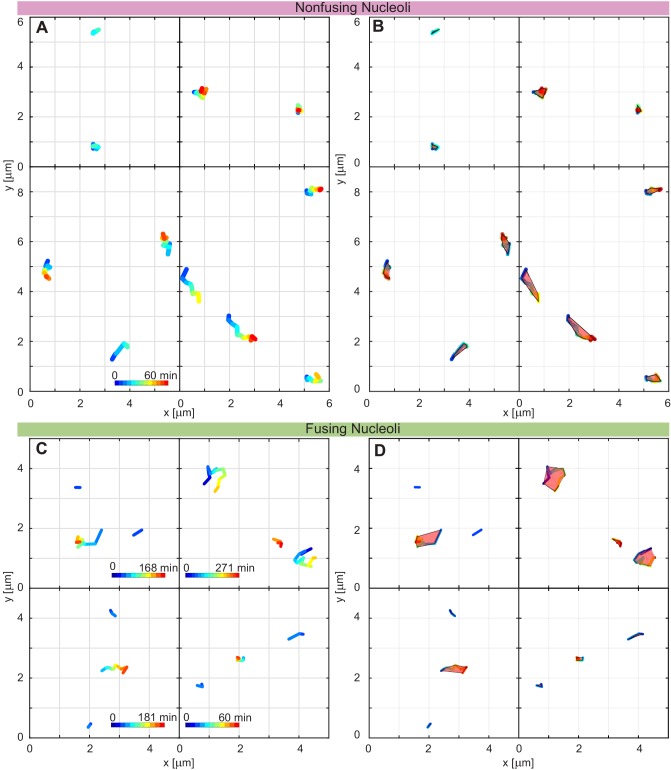
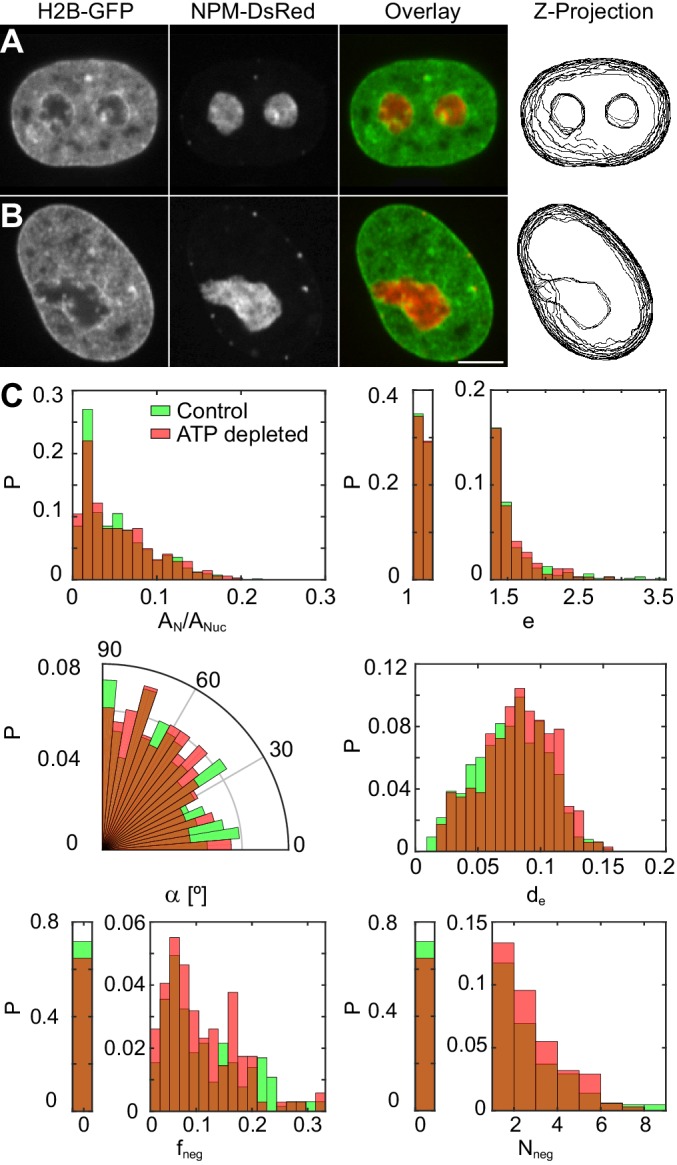
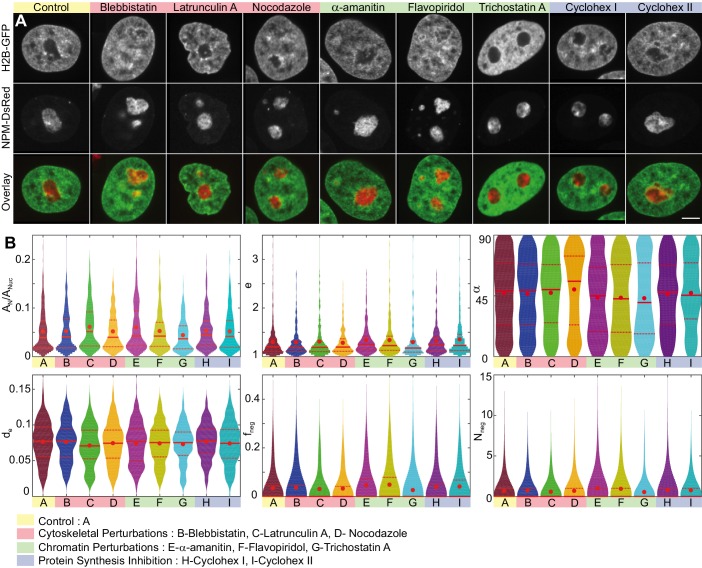
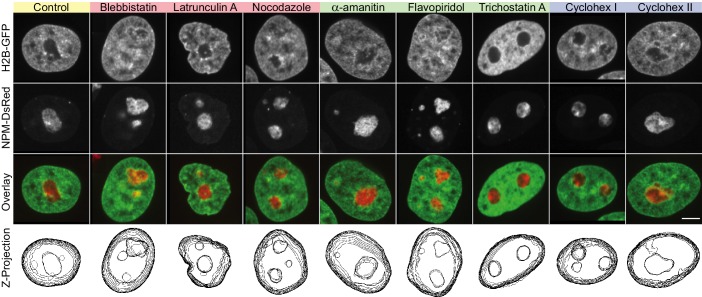
Liquid-liquid phase separation (LLPS) has been recognized as one of the key cellular organizing principles and was shown to be responsible for formation of membrane-less organelles such as nucleoli. Although nucleoli were found to behave like liquid droplets, many ramifications of LLPS including nucleolar dynamics and interactions with the surrounding liquid remain to be revealed. Here, we study the motion of human nucleoli in vivo, while monitoring the shape of the nucleolus-nucleoplasm interface. We reveal two types of nucleolar pair dynamics: an unexpected correlated motion prior to coalescence and an independent motion otherwise. This surprising kinetics leads to a nucleolar volume distribution, [Formula: see text], unaccounted for by any current theory. Moreover, we find that nucleolus-nucleoplasm interface is maintained by ATP-dependent processes and susceptible to changes in chromatin transcription and packing. Our results extend and enrich the LLPS framework by showing the impact of the surrounding nucleoplasm on nucleoli in living cells.
Keywords: human; liquid-liquid phase separation; molecular biophysics; nucleolar dynamics; nucleolus; nucleoplasm; nucleus; physics of living systems; structural biology.
© 2019, Caragine et al.
Conflict of interest statement
CC, SH, AZ No competing interests declared
Figures
References
-
- Alberts B, Johnson A, Lewis J, Morgan D, Raff M, Roberts K, Walter P. Molecular Biology of the Cell. Garland Science; 2014. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
