

Genome-wide identification and characterization of the GDP-L-galactose phosphorylase gene family in bread wheat
- PMID: 31771507
- PMCID: PMC6878703
- DOI: 10.1186/s12870-019-2123-1
Genome-wide identification and characterization of the GDP-L-galactose phosphorylase gene family in bread wheat
Abstract
Background: Ascorbate is a powerful antioxidant in plants and an essential micronutrient for humans. The GDP-L-galactose phosphorylase (GGP) gene encodes the rate-limiting enzyme of the L-galactose pathway-the dominant ascorbate biosynthetic pathway in plants-and is a promising gene candidate for increasing ascorbate in crops. In addition to transcriptional regulation, GGP production is regulated at the translational level through an upstream open reading frame (uORF) in the long 5'-untranslated region (5'UTR). The GGP genes have yet to be identified in bread wheat (Triticum aestivum L.), one of the most important food grain sources for humans.
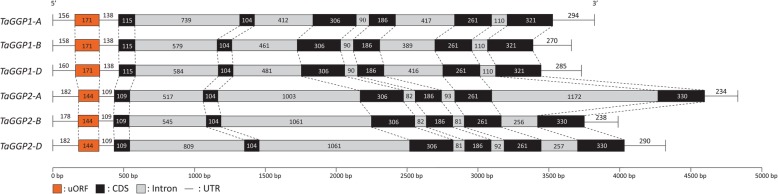
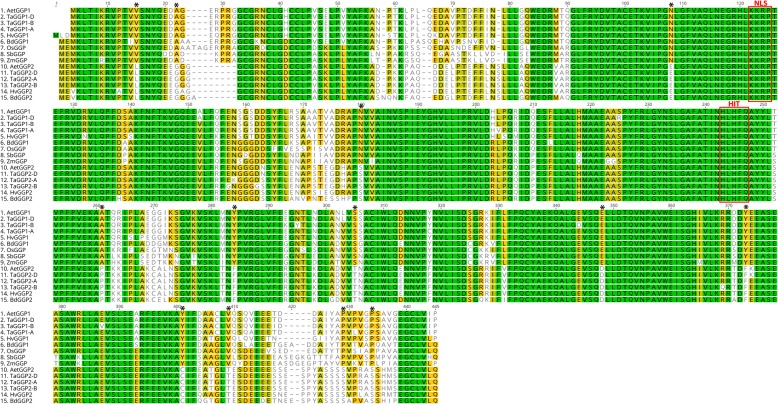
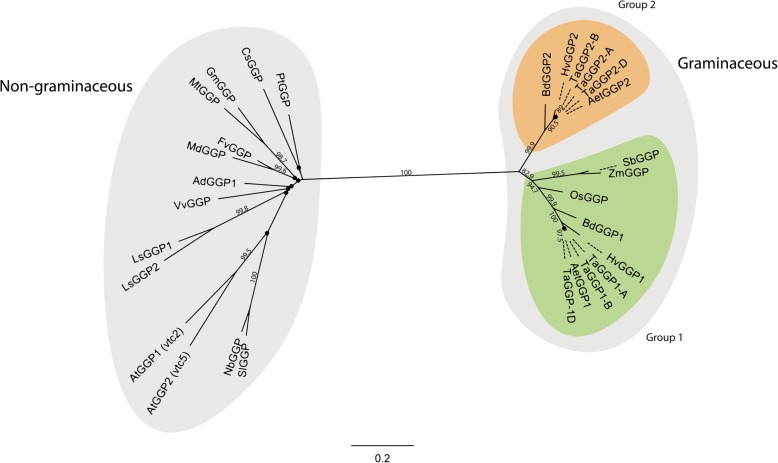
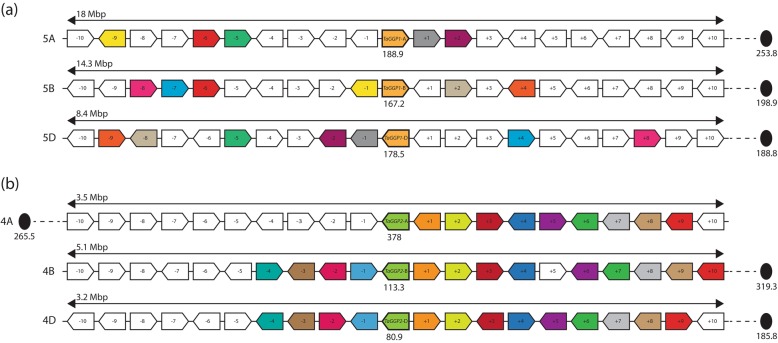
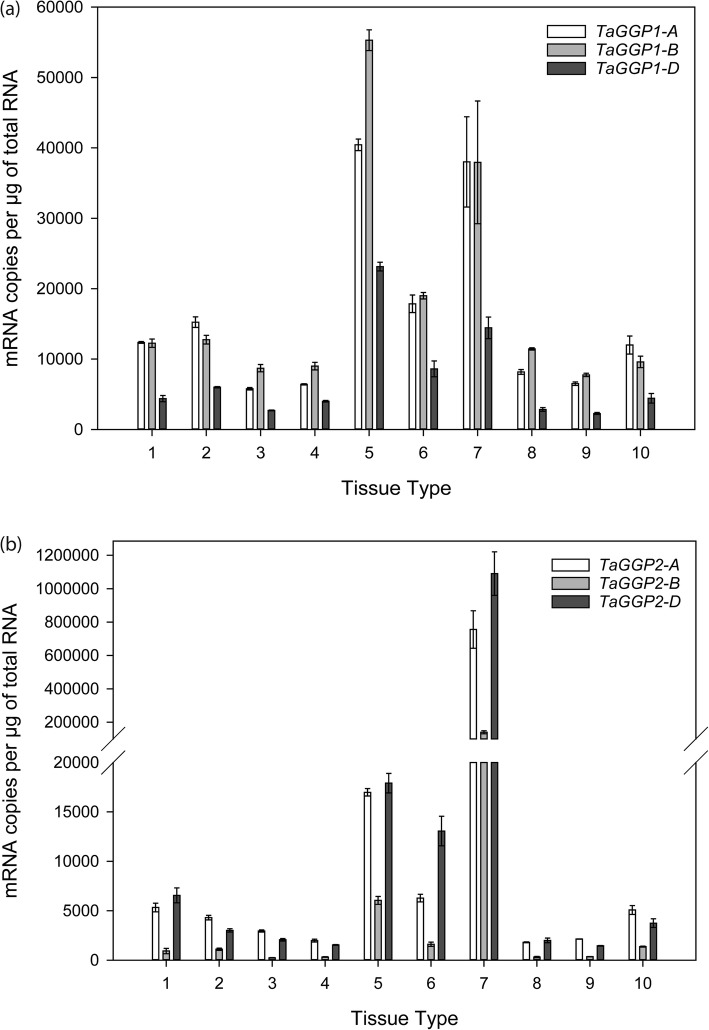
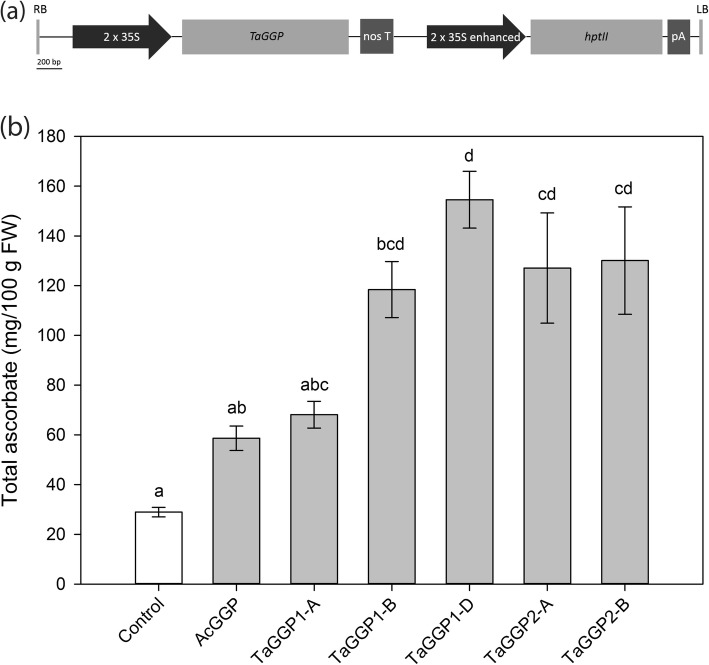
Results: Bread wheat chromosomal groups 4 and 5 were found to each contain three homoeologous TaGGP genes on the A, B, and D subgenomes (TaGGP2-A/B/D and TaGGP1-A/B/D, respectively) and a highly conserved uORF was present in the long 5'UTR of all six genes. Phylogenetic analyses demonstrated that the TaGGP genes separate into two distinct groups and identified a duplication event of the GGP gene in the ancestor of the Brachypodium/Triticeae lineage. A microsynteny analysis revealed that the TaGGP1 and TaGGP2 subchromosomal regions have no shared synteny suggesting that TaGGP2 may have been duplicated via a transposable element. The two groups of TaGGP genes have distinct expression patterns with the TaGGP1 homoeologs broadly expressed across different tissues and developmental stages and the TaGGP2 homoeologs highly expressed in anthers. Transient transformation of the TaGGP coding sequences in Nicotiana benthamiana leaf tissue increased ascorbate concentrations more than five-fold, confirming their functional role in ascorbate biosynthesis in planta.
Conclusions: We have identified six TaGGP genes in the bread wheat genome, each with a highly conserved uORF. Phylogenetic and microsynteny analyses highlight that a transposable element may have been responsible for the duplication and specialized expression of GGP2 in anthers in the Brachypodium/Triticeae lineage. Transient transformation of the TaGGP coding sequences in N. benthamiana demonstrated their activity in planta. The six TaGGP genes and uORFs identified in this study provide a valuable genetic resource for increasing ascorbate concentrations in bread wheat.
Keywords: Ascorbic acid; Gene expression; Phylogeny; Synteny; Transient expression; Upstream open reading frame; Vitamin C.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

Similar articles
-
Enhancing ascorbate in fruits and tubers through over-expression of the L-galactose pathway gene GDP-L-galactose phosphorylase.Plant Biotechnol J. 2012 May;10(4):390-7. doi: 10.1111/j.1467-7652.2011.00668.x. Epub 2011 Dec 1. Plant Biotechnol J. 2012. PMID: 22129455
-
An upstream open reading frame is essential for feedback regulation of ascorbate biosynthesis in Arabidopsis.Plant Cell. 2015 Mar;27(3):772-86. doi: 10.1105/tpc.114.133777. Epub 2015 Feb 27. Plant Cell. 2015. PMID: 25724639 Free PMC article.
-
Comparison of ascorbic acid biosynthesis in different tissues of three non-heading Chinese cabbage cultivars.Plant Physiol Biochem. 2013 Dec;73:229-36. doi: 10.1016/j.plaphy.2013.10.005. Epub 2013 Oct 10. Plant Physiol Biochem. 2013. PMID: 24157701
-
How does light facilitate vitamin C biosynthesis in leaves?Biosci Biotechnol Biochem. 2022 Aug 24;86(9):1173-1182. doi: 10.1093/bbb/zbac096. Biosci Biotechnol Biochem. 2022. PMID: 35746883 Review.
-
The regulation of ascorbate biosynthesis.Curr Opin Plant Biol. 2016 Oct;33:15-22. doi: 10.1016/j.pbi.2016.04.010. Epub 2016 May 11. Curr Opin Plant Biol. 2016. PMID: 27179323 Review.
Cited by
-
Discovery of a conserved translationally repressive upstream open reading frame within the iron-deficiency response regulator IDEF2.BMC Plant Biol. 2024 Sep 30;24(1):891. doi: 10.1186/s12870-024-05473-y. BMC Plant Biol. 2024. PMID: 39343926 Free PMC article.
-
Gene expression profiling in Rosa roxburghii fruit and overexpressing RrGGP2 in tobacco and tomato indicates the key control point of AsA biosynthesis.Front Plant Sci. 2023 Jan 10;13:1096493. doi: 10.3389/fpls.2022.1096493. eCollection 2022. Front Plant Sci. 2023. PMID: 36704162 Free PMC article.
-
Metabolic engineering in food crops to enhance ascorbic acid production: crop biofortification perspectives for human health.Physiol Mol Biol Plants. 2022 Apr;28(4):871-884. doi: 10.1007/s12298-022-01172-w. Epub 2022 Apr 19. Physiol Mol Biol Plants. 2022. PMID: 35464783 Free PMC article. Review.
-
Characterization of the GGP gene family in potato (Solanum tuberosum L.) and Pepper (Capsicum annuum L.) and its expression analysis under hormonal and abiotic stresses.Sci Rep. 2024 Jul 3;14(1):15329. doi: 10.1038/s41598-024-66337-x. Sci Rep. 2024. PMID: 38961199 Free PMC article.
-
Expression Pattern and Functional Analyses of Arabidopsis Guard Cell-Enriched GDSL Lipases.Front Plant Sci. 2021 Sep 21;12:748543. doi: 10.3389/fpls.2021.748543. eCollection 2021. Front Plant Sci. 2021. PMID: 34621289 Free PMC article.
References
-
- Lisko Katherine A., Aboobucker Siddique I., Torres Raquel, Lorence Argelia. Phytochemicals – Biosynthesis, Function and Application. Cham: Springer International Publishing; 2014. Engineering Elevated Vitamin C in Plants to Improve their Nutritional Content, Growth, and Tolerance to Abiotic Stress; pp. 109–128.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
