

A critical role for microglia in maintaining vascular integrity in the hypoxic spinal cord
- PMID: 31772011
- PMCID: PMC6925983
- DOI: 10.1073/pnas.1912178116
A critical role for microglia in maintaining vascular integrity in the hypoxic spinal cord
Abstract
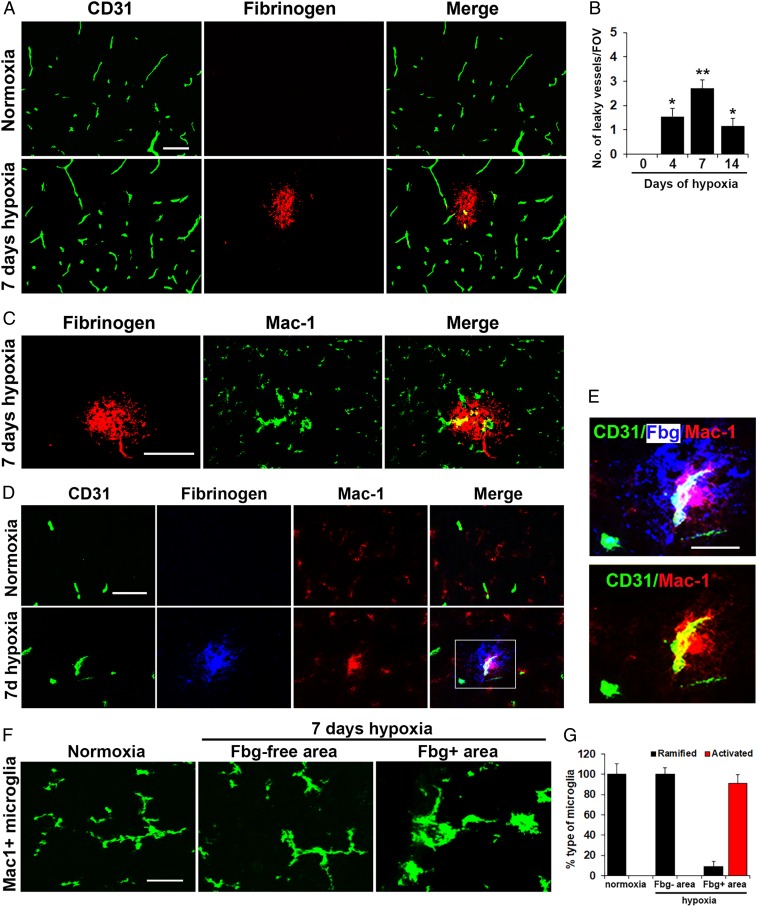
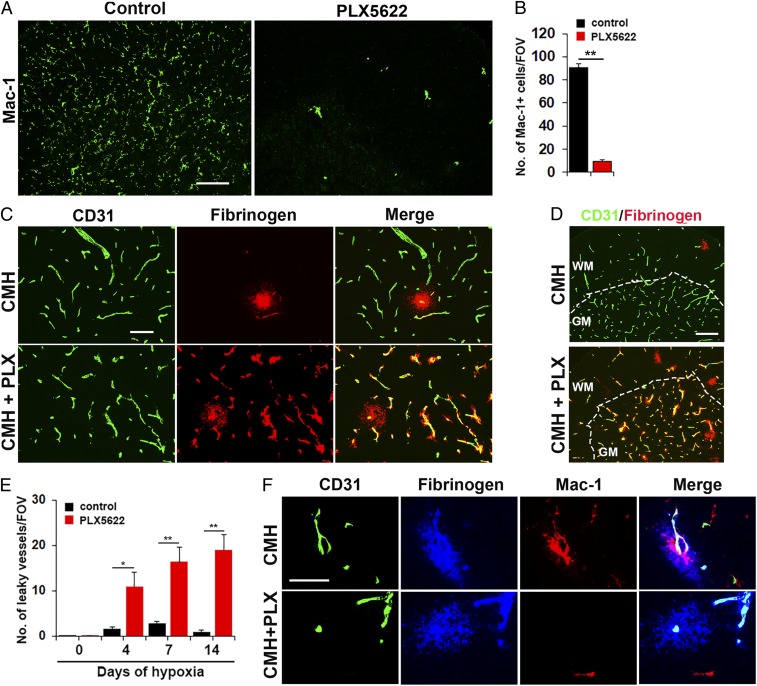
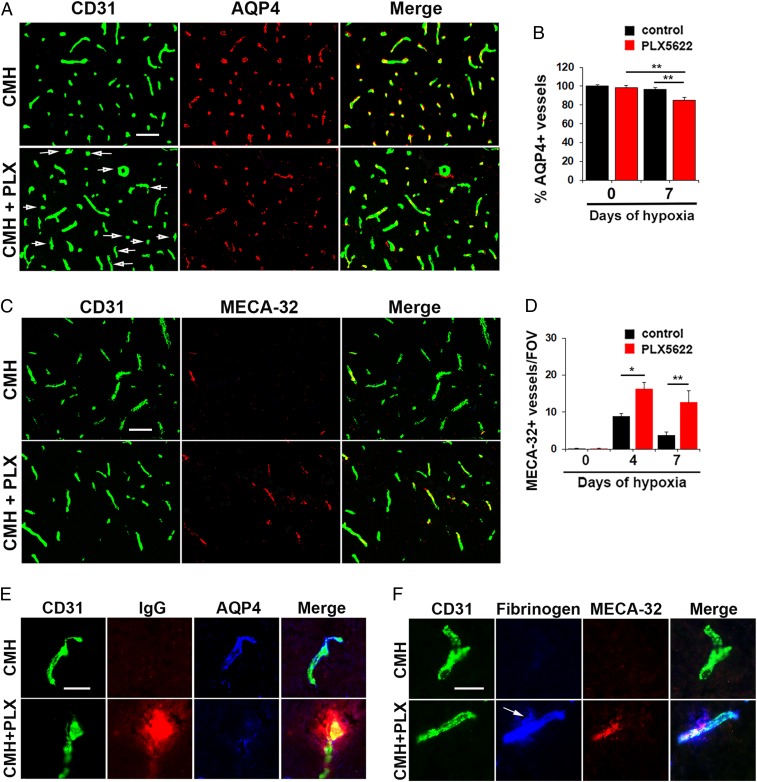
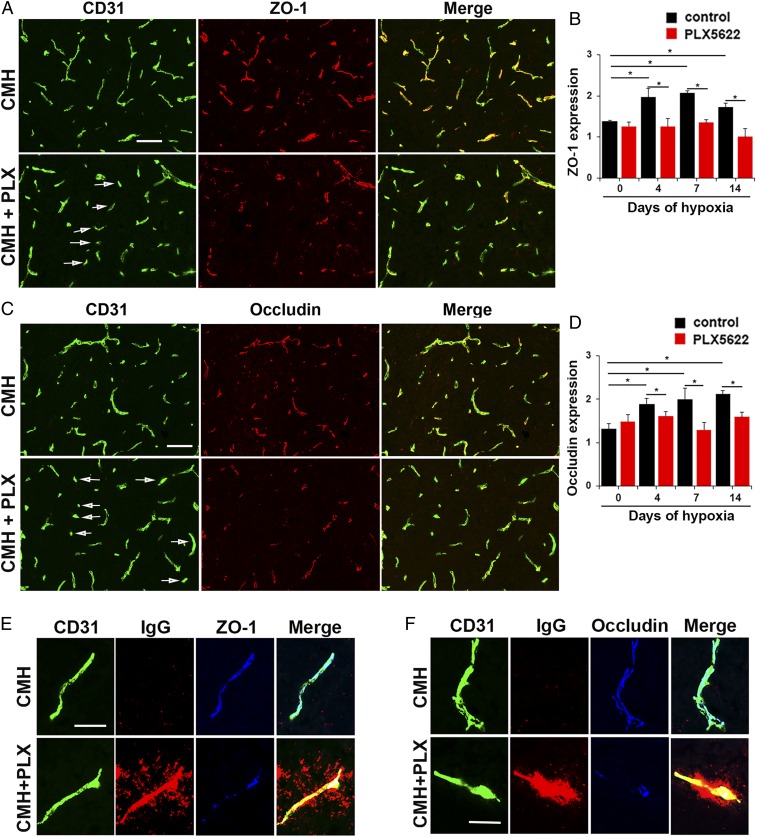
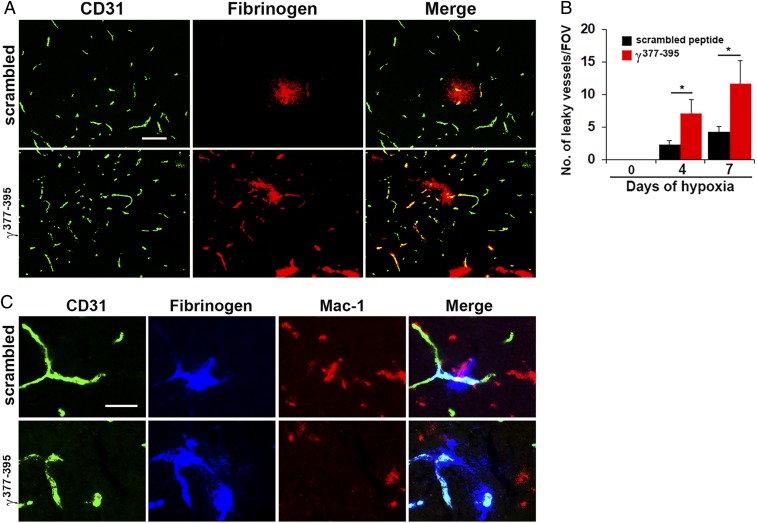
Hypoxic preconditioning reduces disease severity in a mouse model of multiple sclerosis (MS), in part by enhancing the barrier properties of spinal cord blood vessels. Because other studies have shown that similar levels of hypoxia transiently increase permeability of central nervous system (CNS) blood vessels, the goal of this study was to define the impact of chronic mild hypoxia (CMH, 8% O2) on the integrity of spinal cord blood vessels and the responses of neighboring glial cells. Using extravascular fibrinogen as a marker of vascular disruption, we found that CMH triggered transient vascular leak in spinal cord blood vessels, particularly in white matter, which was associated with clustering and activation of Mac-1-positive microglia around disrupted vessels. Microglial depletion with the colony stimulating factor-1 receptor (CSF-1R) inhibitor PLX5622, while having no effect under normoxic conditions, profoundly increased vascular leak in both white and gray matter during CMH, and this was associated with disruption of astrocyte-vascular coupling and enhanced loss of tight junction proteins. Microglial repair of leaky blood vessels was blocked by a peptide that inhibits the interaction between fibrinogen and its Mac-1 integrin receptor. These findings highlight an important role for microglia in maintaining vascular integrity in the hypoxic spinal cord and suggest that a fibrinogen-Mac-1 interaction underpins this response. As relative hypoxia is experienced in many situations including high altitude, lung disease, obstructive sleep apnea, and age-related CNS ischemia/hypoxia, our findings have important implications regarding the critical role of microglia in maintaining vascular integrity in the CNS.
Keywords: blood vessels; fibrinogen; hypoxia; microglia; spinal cord.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Ballabh P., Braun A., Nedergaard M., The blood-brain barrier: An overview: Structure, regulation, and clinical implications. Neurobiol. Dis. 16, 1–13 (2004). - PubMed
-
- Huber J. D., Egleton R. D., Davis T. P., Molecular physiology and pathophysiology of tight junctions in the blood-brain barrier. Trends Neurosci. 24, 719–725 (2001). - PubMed
-
- Pardridge W. M., Blood-brain barrier drug targeting: The future of brain drug development. Mol. Interv. 3, 90–105 (2003). - PubMed
-
- del Zoppo G. J., Milner R., Integrin-matrix interactions in the cerebral microvasculature. Arterioscler. Thromb. Vasc. Biol. 26, 1966–1975 (2006). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
