

Cicadidae Periostracum, the Cast-Off Skin of Cicada, Protects Dopaminergic Neurons in a Model of Parkinson's Disease
- PMID: 31772707
- PMCID: PMC6854990
- DOI: 10.1155/2019/5797512
Cicadidae Periostracum, the Cast-Off Skin of Cicada, Protects Dopaminergic Neurons in a Model of Parkinson's Disease
Abstract
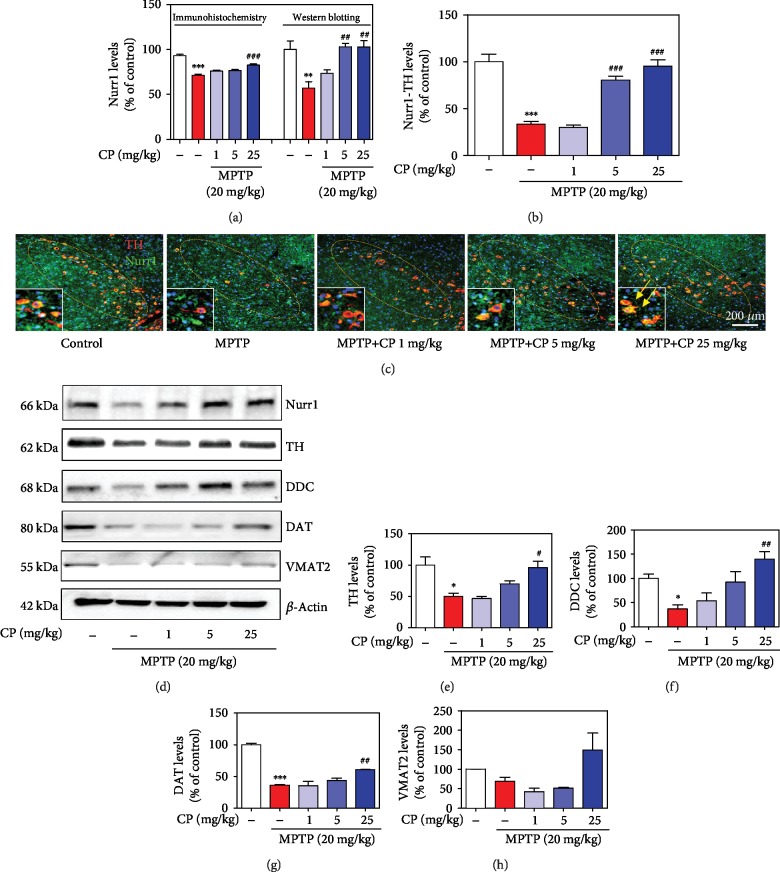
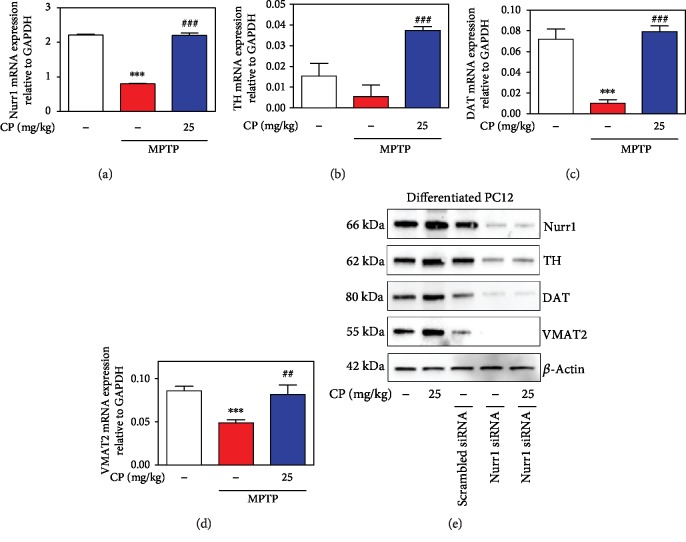
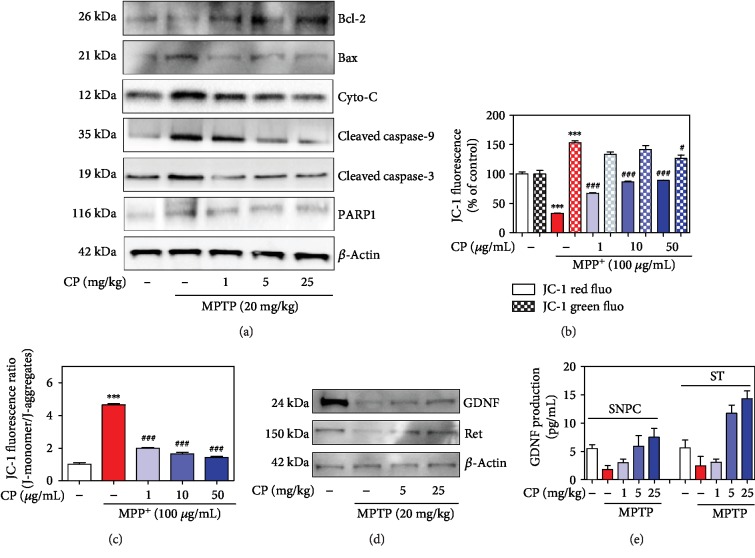
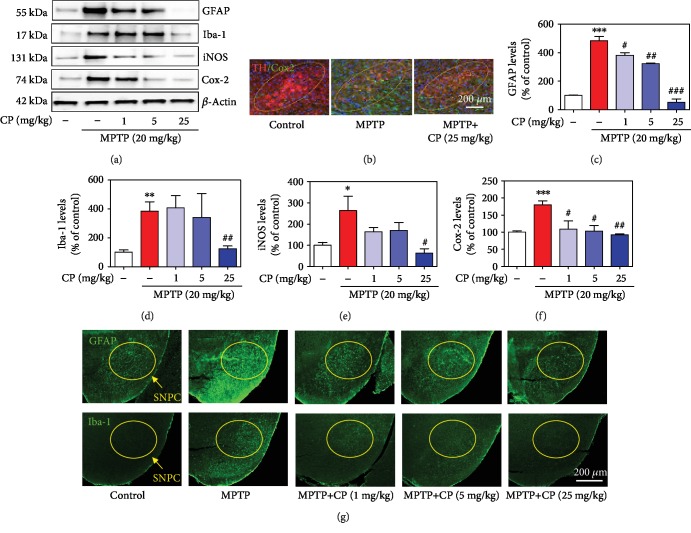
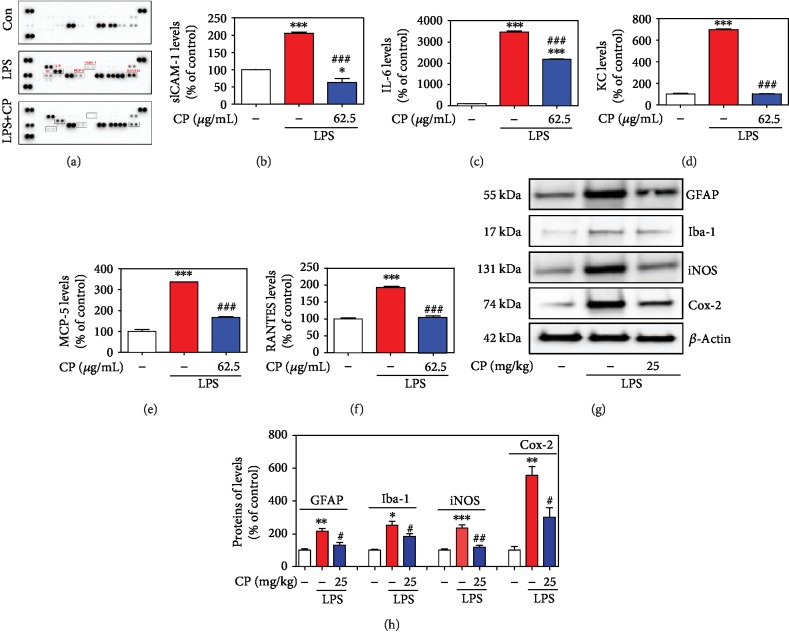
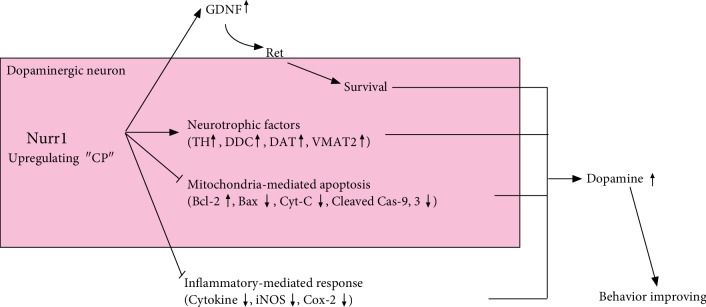
Parkinson's disease (PD) is characterized by dopaminergic neuronal loss in the substantia nigra pars compacta (SNPC) and the striatum. Nuclear receptor-related 1 protein (Nurr1) is a nuclear hormone receptor implicated in limiting mitochondrial dysfunction, apoptosis, and inflammation in the central nervous system and protecting dopaminergic neurons and a promising therapeutic target for PD. Cicadidae Periostracum (CP), the cast-off skin of Cryptotympana pustulata Fabricius, has been used in traditional medicine for its many clinical pharmacological effects, including the treatment of psychological symptoms in PD. However, scientific evidence for the use of CP in neurodegenerative diseases, including PD, is lacking. Here, we investigated the protective effects of CP on 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine- (MPTP-) induced PD in mice and explored the underlying mechanisms of action, focusing on Nurr1. CP increased the expression levels of Nurr1, tyrosine hydroxylase, DOPA decarboxylase, dopamine transporter, and vesicular monoamine transporter 2 via extracellular signal-regulated kinase phosphorylation in differentiated PC12 cells and the mouse SNPC. In MPTP-induced PD, CP promoted recovery from movement impairments. CP prevented dopamine depletion and protected against dopaminergic neuronal degradation via mitochondria-mediated apoptotic proteins such as B-cell lymphoma 2 (Bcl-2), Bcl-2-associated X, cytochrome c, and cleaved caspase-9 and caspase-3 by inhibiting MPTP-induced neuroinflammatory cytokines, inducible nitric oxide synthase, cyclooxygenase 2, and glial/microglial activation. Moreover, CP inhibited lipopolysaccharide-induced neuroinflammatory cytokines and response levels and glial/microglial activation in BV2 microglia and the mouse brain. Our findings suggest that CP might contribute to neuroprotective signaling by regulating neurotrophic factors primarily via Nurr1 signaling, neuroinflammation, and mitochondria-mediated apoptosis.
Copyright © 2019 Hye-Sun Lim et al.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
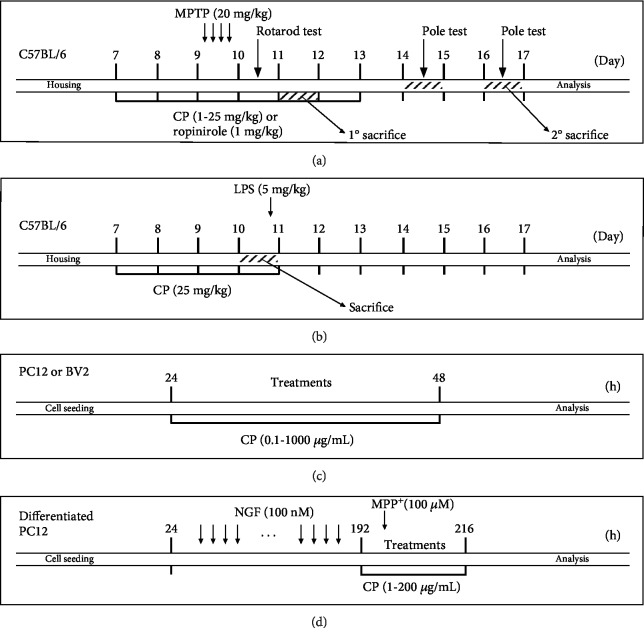
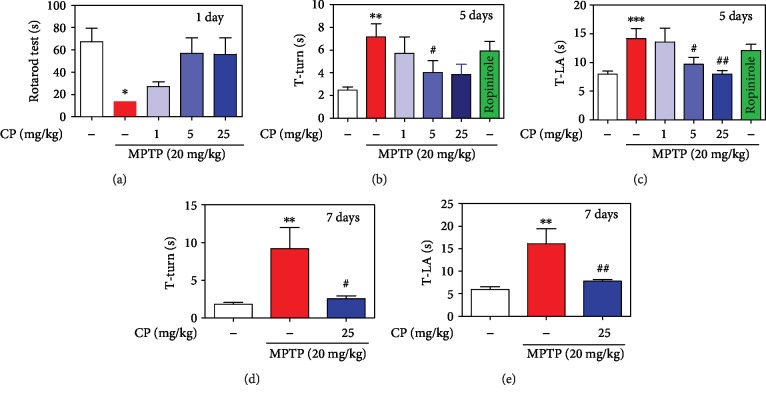
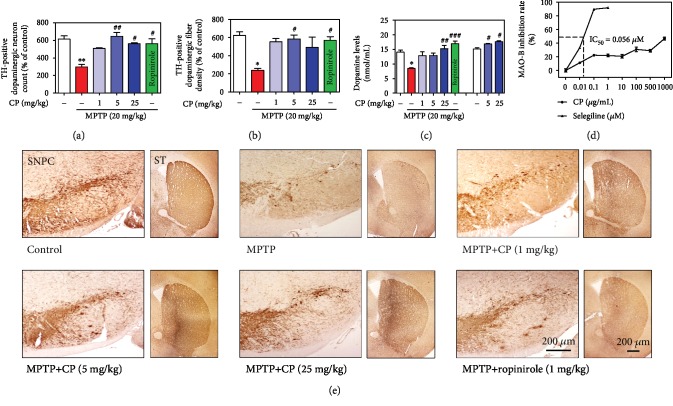
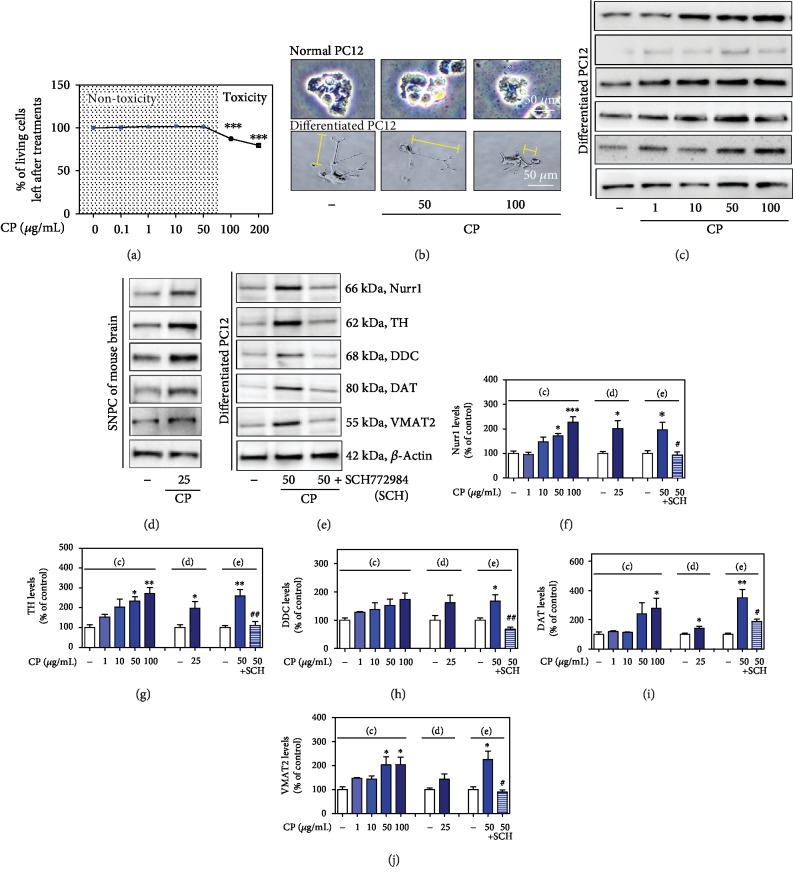
References
-
- Kim C.-H., Han B.-S., Moon J., et al. Nuclear receptor Nurr1 agonists enhance its dual functions and improve behavioral deficits in an animal model of Parkinson’s disease. Proceedings of the National Academy of Sciences of the United States of America. 2015;112(28):8756–8761. doi: 10.1073/pnas.1509742112. - DOI - PMC - PubMed
-
- Dong J., Wang Y., Liu X., Le W. Nurr1 deficiency-mediated inflammatory injury to nigral dopamine neurons in Parkinson’s disease. Parkinsonism & Related Disorders. 2018;46, article e66 doi: 10.1016/j.parkreldis.2017.11.226. - DOI
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
