

Hypoxia-induced autophagy drives colorectal cancer initiation and progression by activating the PRKC/PKC-EZR (ezrin) pathway
- PMID: 31775562
- PMCID: PMC7469473
- DOI: 10.1080/15548627.2019.1687213
Hypoxia-induced autophagy drives colorectal cancer initiation and progression by activating the PRKC/PKC-EZR (ezrin) pathway
Abstract
In solid tumors, cancer stem cells (CSCs) or tumor-initiating cells (TICs) are often found in hypoxic niches. Nevertheless, the influence of hypoxia on TICs is poorly understood. Using previously established, TIC-enrichedpatient-derived colorectal cancer (CRC) cultures, we show that hypoxia increases the self-renewal capacity of TICs while inducing proliferation arrest in their more differentiated counterpart cultures. Gene expression data revealed macroautophagy/autophagy as one of the major pathways induced by hypoxia in TICs. Interestingly, hypoxia-induced autophagy was found to induce phosphorylation of EZR (ezrin) at Thr567 residue, which could be reversed by knocking down ATG5, BNIP3, BNIP3L, or BECN1. Furthermore, we identified PRKCA/PKCα as a potential kinase involved in hypoxia-induced autophagy-mediated TIC self-renewal. Genetic targeting of autophagy or pharmacological inhibition of PRKC/PKC and EZR resulted in decreased tumor-initiating potential of TICs. In addition, we observed significantly reduced in vivo tumor initiation and growth after a stable knockdown of ATG5. Analysis of human CRC samples showed that p-EZR is often present in TICs located in the hypoxic and autophagic regions of the tumor. Altogether, our results establish the hypoxia-autophagy-PKC-EZR signaling axis as a novel regulatory mechanism of TIC self-renewal and CRC progression. Autophagy inhibition might thus represent a promising therapeutic strategy for cancer patients.
Abbreviations: ATG: autophagy related; BECN1: beclin 1; BNIP3: BCL2 interacting protein 3; BNIP3L: BCL2 interacting protein 3 like; CQ: chloroquine; CSC: cancer stem cells; CRC: colorectal cancer; HIF1A/HIF-1α: hypoxia inducible factor 1 subunit alpha; MAP1LC3/LC3: microtubule associated protein 1 light chain 3; PRKC/PKC: protein kinase C; SQSTM1/p62: sequestosome 1; TICs: tumor-initiating cells.
Keywords: Autophagy; cancer stem cell; colorectal cancer; ezrin; hypoxia; protein kinase C; self-renewal capacity; tumor-initiating cell.
Conflict of interest statement
No potential conflict of interest was reported by the authors.
Figures

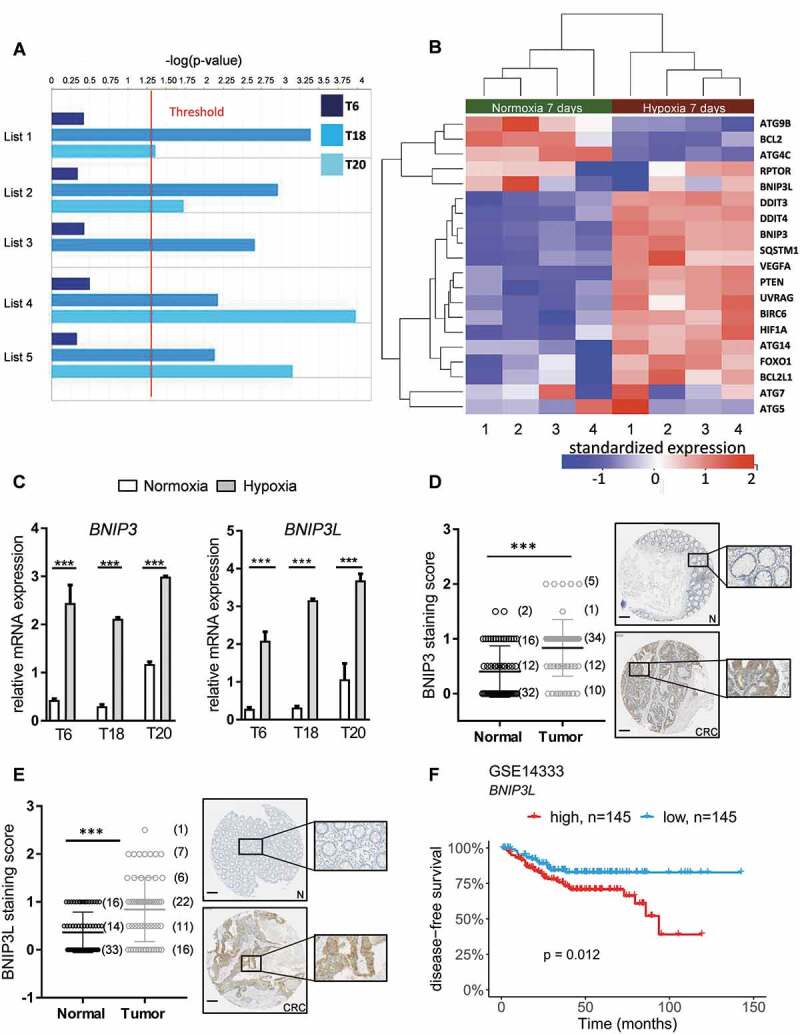


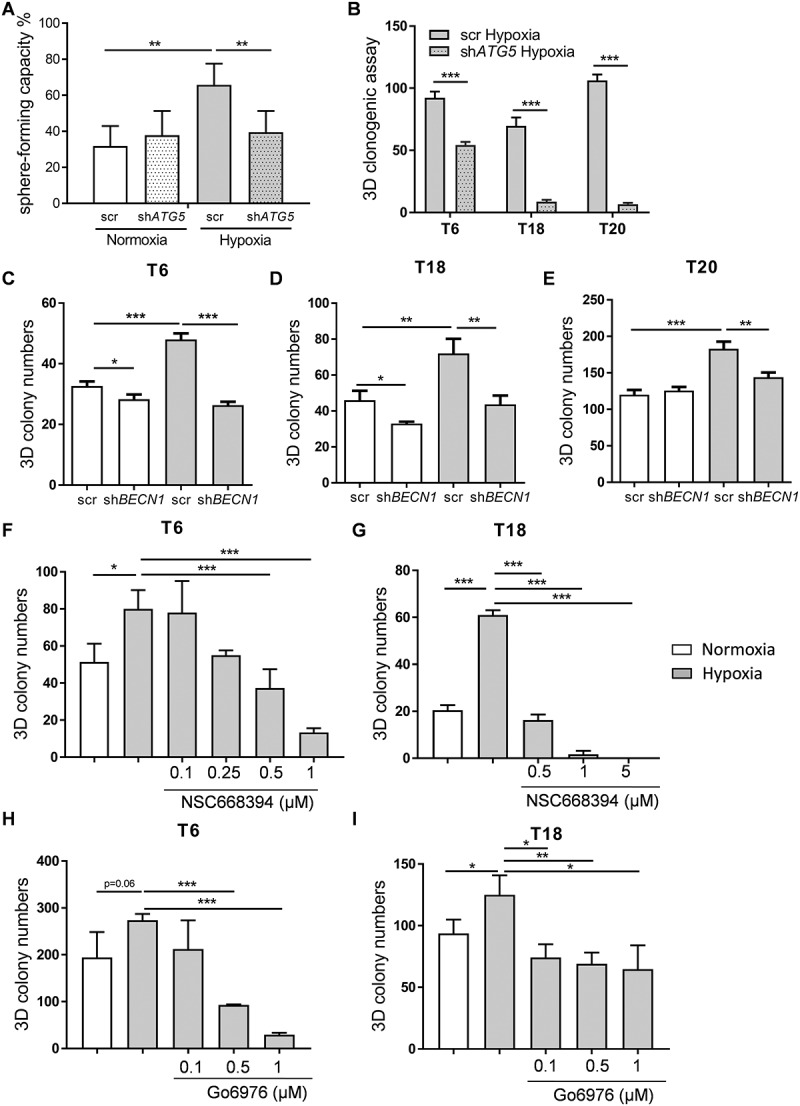


References
-
- Siegel RL, Miller KD, Jemal A.. Cancer statistics, 2016. CA Cancer J Clin [Internet]. 2016. [cited 2016 September16]; 66:7–30. Available from: http://www.ncbi.nlm.nih.gov/pubmed/26742998 - PubMed
-
- Qureshi-Baig K, Ullmann P, Haan S, et al. Tumor-initiating cells: a criTICal review of isolation approaches and new challenges in targeting strategies. Mol Cancer [Internet]. 2017. [cited 2017 April27]; 16:40. Available from: http://molecular-cancer.biomedcentral.com/articles/10.1186/s12943-017-06... - DOI - PMC - PubMed
-
- Schito L, Semenza GL.. Hypoxia-inducible factors: master regulators of cancer progression. Trends Cancer [Internet]. 2016. [cited 2018 August22]; 2:758–770. Available from: http://www.ncbi.nlm.nih.gov/pubmed/28741521 - PubMed
-
- Carnero A, Lleonart M. The hypoxic microenvironment: a determinant of cancer stem cell evolution. BioEssays [Internet]. 2016. [cited 2016 November2]; 38:S65–74. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous