

The effects of n-6 polyunsaturated fatty acid deprivation on the inflammatory gene response to lipopolysaccharide in the mouse hippocampus
- PMID: 31775778
- PMCID: PMC6882015
- DOI: 10.1186/s12974-019-1615-0
The effects of n-6 polyunsaturated fatty acid deprivation on the inflammatory gene response to lipopolysaccharide in the mouse hippocampus
Abstract
Background: Neuroinflammation is thought to contribute to psychiatric and neurological disorders such as major depression and Alzheimer's disease (AD). N-6 polyunsaturated fatty acids (PUFA) and molecules derived from them, including linoleic acid- and arachidonic acid-derived lipid mediators, are known to have pro-inflammatory properties in the periphery; however, this has yet to be tested in the brain. Lowering the consumption of n-6 PUFA is associated with a decreased risk of depression and AD in human observational studies. The purpose of this study was to investigate the inflammation-modulating effects of lowering dietary n-6 PUFA in the mouse hippocampus.
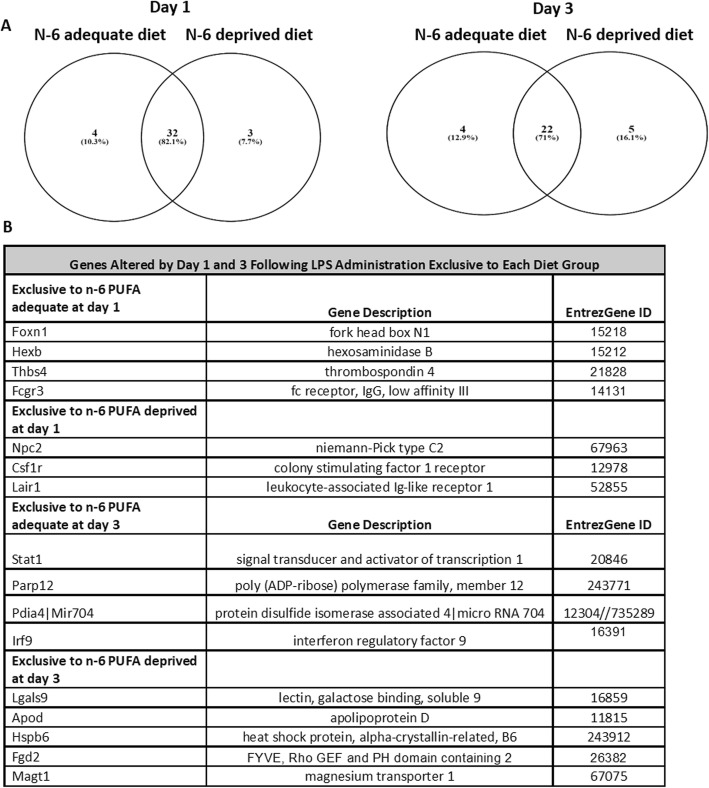
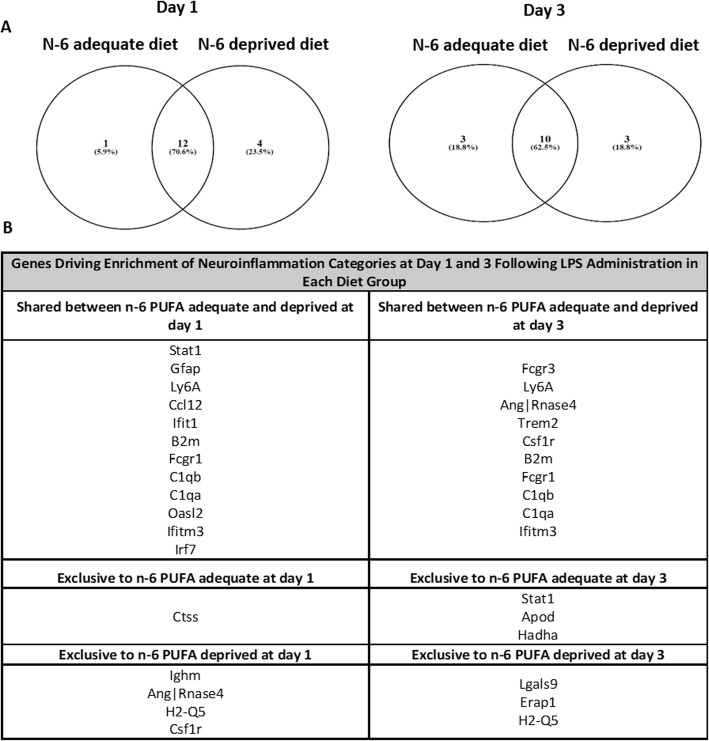
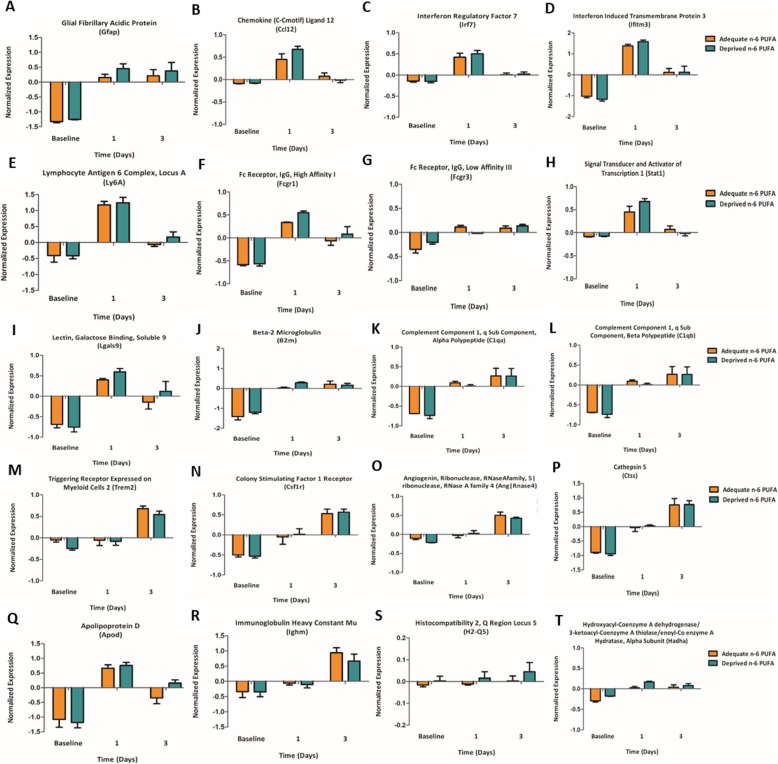
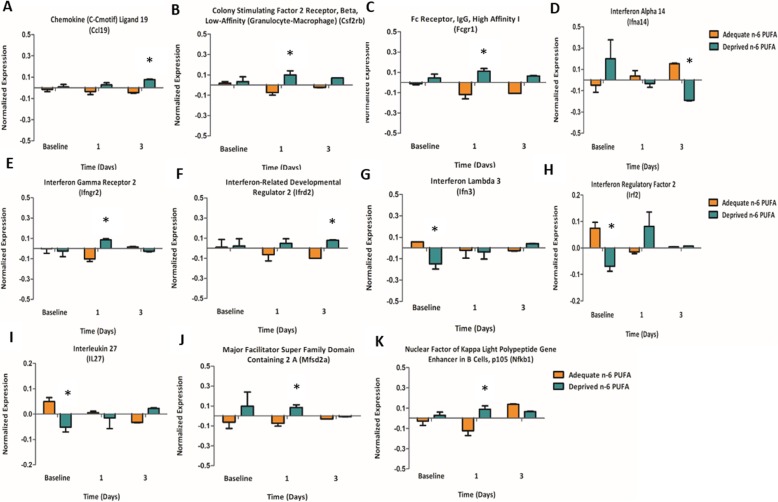
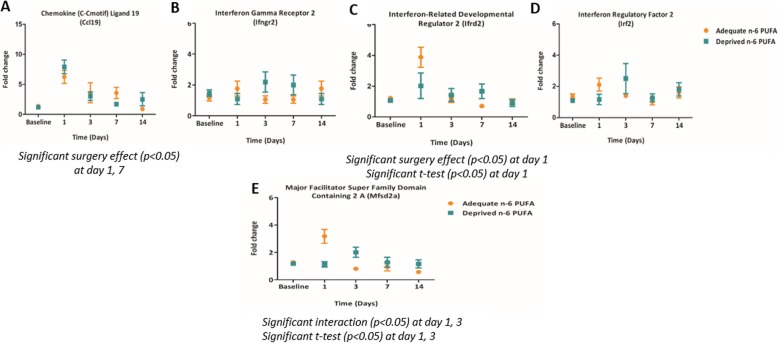
Methods: C57BL/6 male mice were fed either an n-6 PUFA deprived (2% of total fatty acids) or an n-6 PUFA adequate (23% of total fatty acids) diet from weaning to 12 weeks of age. Animals then underwent intracerebroventricular surgery, in which lipopolysaccharide (LPS) was injected into the left lateral ventricle of the brain. Hippocampi were collected at baseline and following LPS administration (1, 3, 7, and 14 days). A microarray (n = 3 per group) was used to identify candidate genes and results were validated by real-time PCR in a separate cohort of animals (n = 5-8 per group).
Results: Mice administered with LPS had significantly increased Gene Ontology categories associated with inflammation and immune responses. These effects were independent of changes in gene expression in any diet group. Results were validated for the effect of LPS treatment on astrocyte, cytokine, and chemokine markers, as well as some results of the diets on Ifrd2 and Mfsd2a expression.
Conclusions: LPS administration increases pro-inflammatory and lipid-metabolizing gene expression in the mouse hippocampus. An n-6 PUFA deprived diet modulated inflammatory gene expression by both increasing and decreasing inflammatory gene expression, without impairing the resolution of neuroinflammation following LPS administration.
Keywords: Arachidonic acid; Hippocampus; Linoleic acid; Lipopolysaccharide; N-6 polyunsaturated fatty acids; Neuroinflammation; mRNA.
Conflict of interest statement
RPB has received research grants from Bunge Ltd., Arctic Nutrition, the Dairy Farmers of Canada, and Nestle Inc., as well as travel support from Mead Johnson and mass spectrometry equipment and support from Sciex. RPB is on the executive of the International Society for the Study of Fatty acids and Lipids and held a meeting on behalf of Fatty Acids and Cell Signalling, both of which rely on corporate sponsorship. RPB has given expert testimony in relation to supplements and the brain. RPB also provides complimentary fatty acid analysis for farmers, food producers, and others involved in the food industry, some of whom provide free food samples.
Figures


References
-
- Heneka MT, Carson MJ, Khoury JE, Landreth GE, Brosseron F, Feinstein DL, Jacobs AH, Wyss-Coray T, Vitorica J, Ransohoff RM, Herrup K, Frautschy SA, Finsen B, Brown GC, Verkhratsky A, Yamanaka K, Koistinaho J, Latz E, Halle A, Petzold GC, Town T, Morgan D, Shinohara ML, Perry VH, Holmes C, Bazan NG, Brooks DJ, Hunot S, Joseph B, Deigendesch N, Garaschuk O, Boddeke E, Dinarello CA, Breitner JC, Cole GM, Golenbock DT, Kummer MP. Neuroinflammation in Alzheimer’s disease. Lancet Neurol. 2015;14:388–405. doi: 10.1016/S1474-4422(15)70016-5. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
