

Pneumolysin Induces 12-Lipoxygenase-Dependent Neutrophil Migration during Streptococcus pneumoniae Infection
- PMID: 31776202
- PMCID: PMC7195902
- DOI: 10.4049/jimmunol.1800748
Pneumolysin Induces 12-Lipoxygenase-Dependent Neutrophil Migration during Streptococcus pneumoniae Infection
Abstract
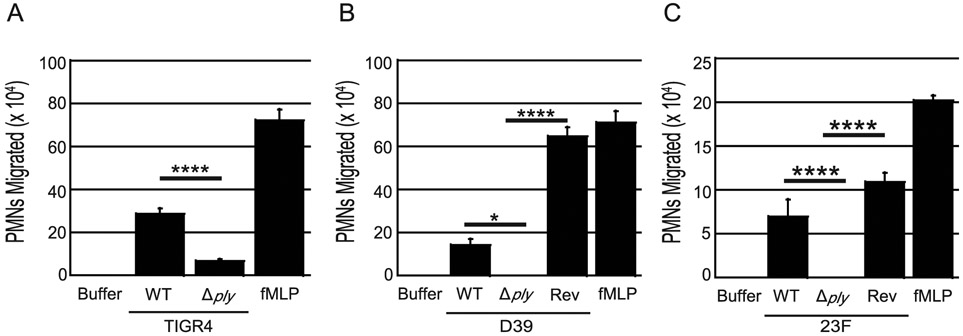
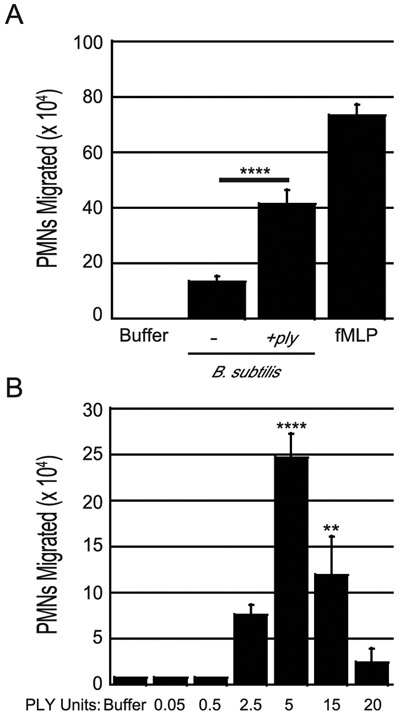
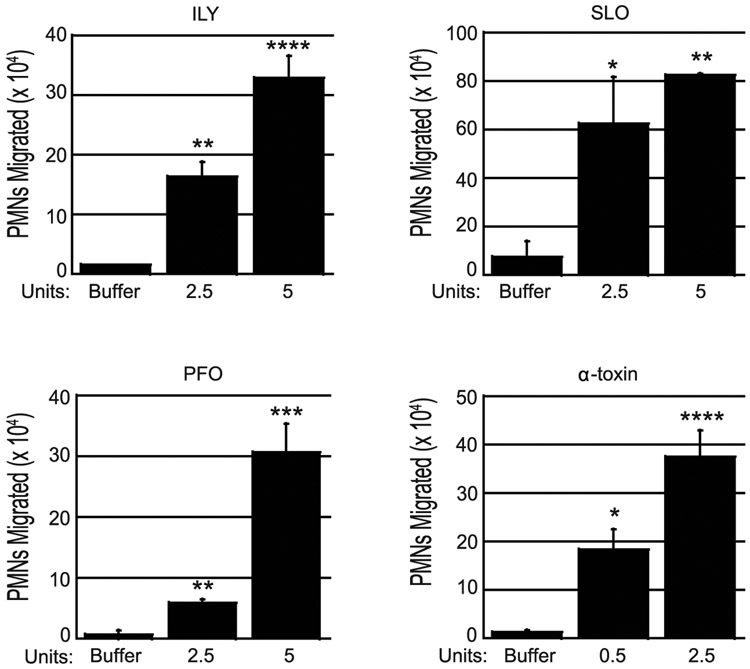
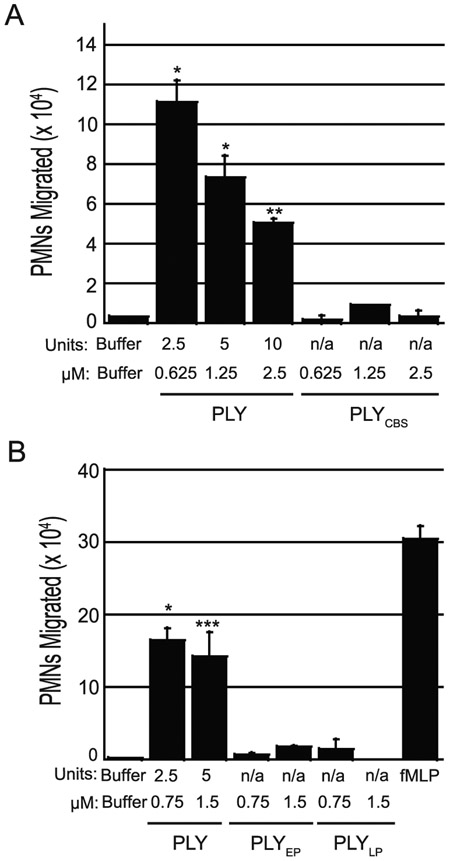
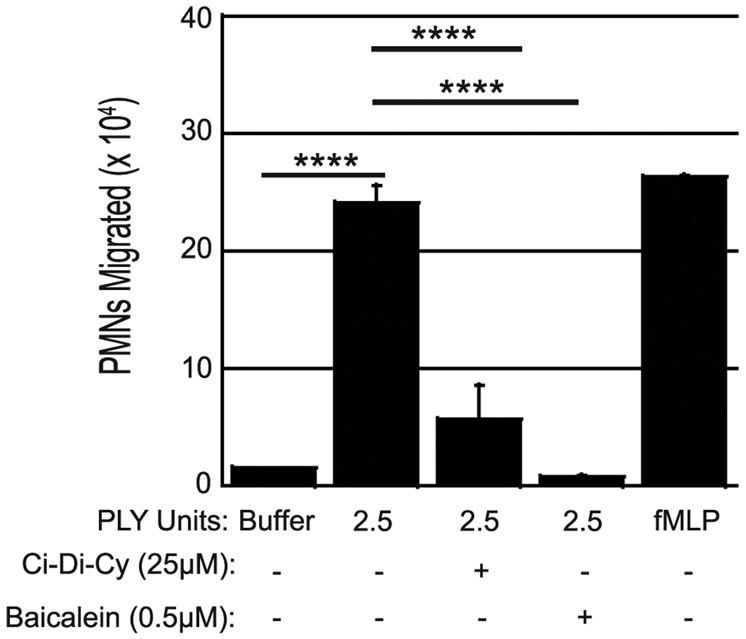
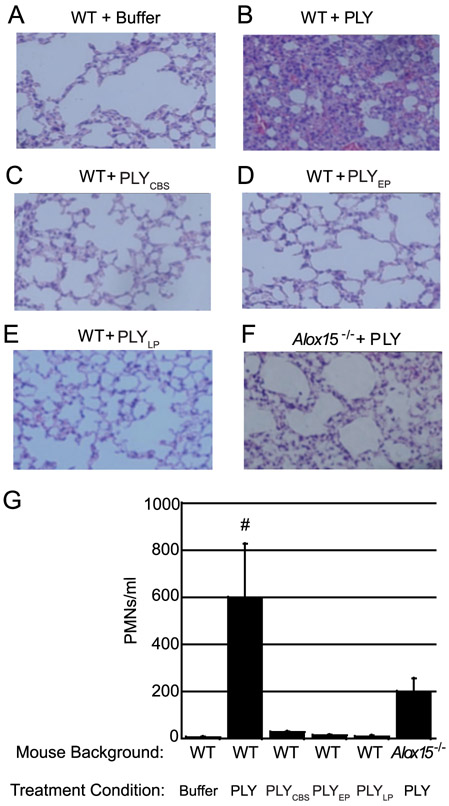
Streptococcus pneumoniae is a major cause of pneumonia, wherein infection of respiratory mucosa drives a robust influx of neutrophils. We have previously shown that S. pneumoniae infection of the respiratory epithelium induces the production of the 12-lipoxygenase (12-LOX)-dependent lipid inflammatory mediator hepoxilin A3, which promotes recruitment of neutrophils into the airways, tissue damage, and lethal septicemia. Pneumolysin (PLY), a member of the cholesterol-dependent cytolysin (CDC) family, is a major S. pneumoniae virulence factor that generates ∼25-nm diameter pores in eukaryotic membranes and promotes acute inflammation, tissue damage, and bacteremia. We show that a PLY-deficient S. pneumoniae mutant was impaired in triggering human neutrophil transepithelial migration in vitro. Ectopic production of PLY endowed the nonpathogenic Bacillus subtilis with the ability to trigger neutrophil recruitment across human-cultured monolayers. Purified PLY, several other CDC family members, and the α-toxin of Clostridium septicum, which generates pores with cross-sectional areas nearly 300 times smaller than CDCs, reproduced this robust neutrophil transmigration. PLY non-pore-forming point mutants that are trapped at various stages of pore assembly did not recruit neutrophils. PLY triggered neutrophil recruitment in a 12-LOX-dependent manner in vitro. Instillation of wild-type PLY but not inactive derivatives into the lungs of mice induced robust 12-LOX-dependent neutrophil migration into the airways, although residual inflammation induced by PLY in 12-LOX-deficient mice indicates that 12-LOX-independent pathways also contribute to PLY-triggered pulmonary inflammation. These data indicate that PLY is an important factor in promoting hepoxilin A3-dependent neutrophil recruitment across pulmonary epithelium in a pore-dependent fashion.
Copyright © 2019 by The American Association of Immunologists, Inc.
Figures
References
-
- Lloyd-Evans N, O’Dempsey TJ, Baldeh I, Secka O, Demba E, Todd JE, Mcardle TF, Banya WS, and Greenwood BM. 1996. Nasopharyngeal carriage of pneumococci in Gambian children and in their families. Pediatr. Infect. Dis. J 15: 866–71. - PubMed
-
- Beckenhaupt P, Bryant P, Kroger A, Weaver D, Wolicki J, W. P 2012. Centers for Disease Control and Prevention Epidemiology and Prevention of Vaccibe-Preventable Diseases Epidemiol. Prev. Vaccine-Preventable Dis 12th Editi: 512.
-
- Huang SS, Johnson KM, Ray GT, Wroe P, Lieu TA, Moore MR, Zell ER, Linder JA, Grijalva CG, Metlay JP, and Finkelstein JA. 2011. Healthcare utilization and cost of pneumococcal disease in the United States. Vaccine 29: 3398–3412. - PubMed
-
- 2013. WHO ∣ Estimated Hib and pneumococcal deaths for children under 5 years of age, 2008. WHO.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
