

BBX4, a phyB-interacting and modulated regulator, directly interacts with PIF3 to fine tune red light-mediated photomorphogenesis
- PMID: 31776262
- PMCID: PMC6925995
- DOI: 10.1073/pnas.1915149116
BBX4, a phyB-interacting and modulated regulator, directly interacts with PIF3 to fine tune red light-mediated photomorphogenesis
Erratum in
-
Correction for Heng et al., BBX4, a phyB-interacting and modulated regulator, directly interacts with PIF3 to fine tune red light-mediated photomorphogenesis.Proc Natl Acad Sci U S A. 2020 Feb 25;117(8):4429-4430. doi: 10.1073/pnas.2001373117. Epub 2020 Feb 18. Proc Natl Acad Sci U S A. 2020. PMID: 32071210 Free PMC article. No abstract available.
Abstract
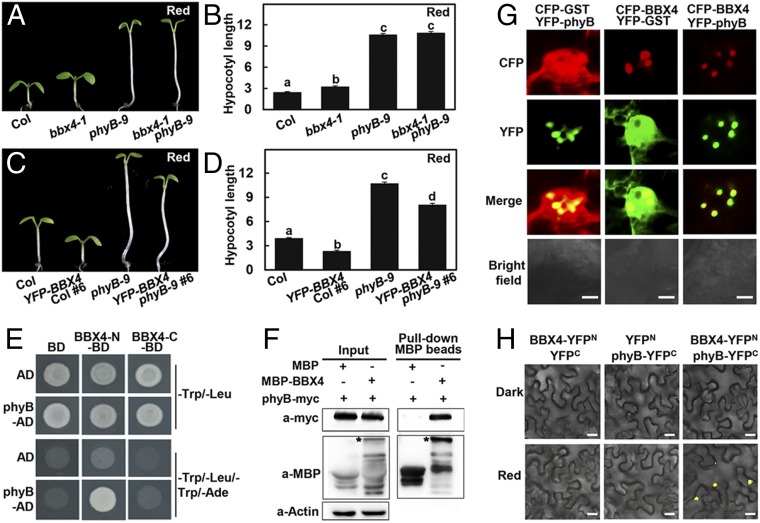
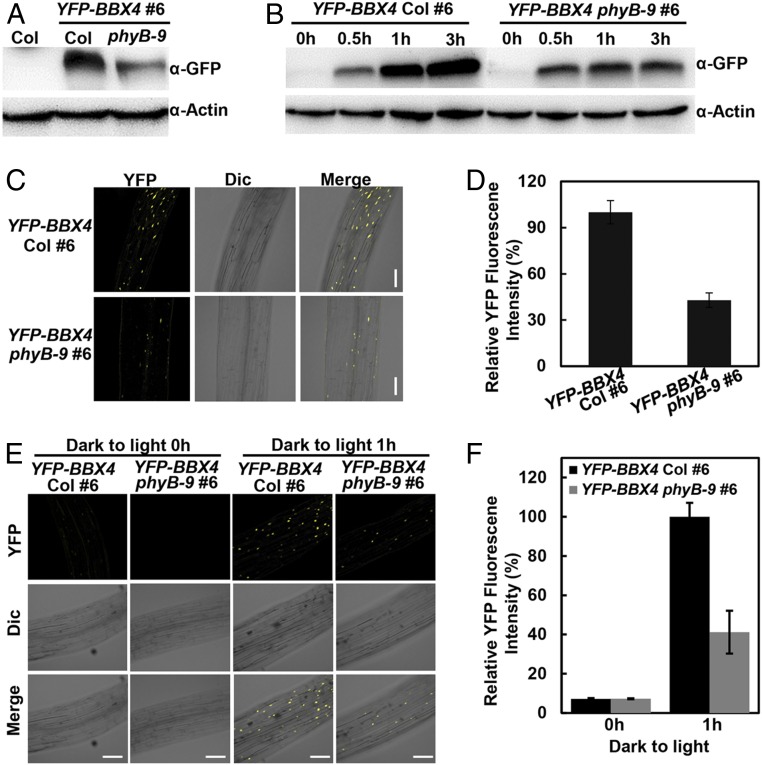
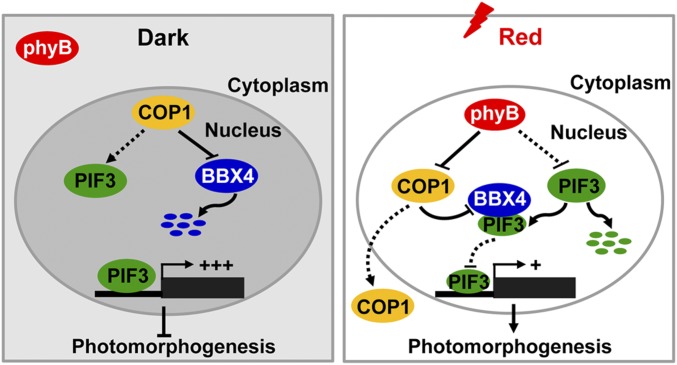
Phytochrome B (phyB) absorbs red light signals and subsequently initiates a set of molecular events in plant cells to promote photomorphogenesis. Here we show that phyB directly interacts with B-BOX CONTAINING PROTEIN 4 (BBX4), a positive regulator of red light signaling, and positively controls its abundance in red light. BBX4 associates with PHYTOCHROME INTERACTING FACTOR 3 (PIF3) and represses PIF3 transcriptional activation activity and PIF3-controlled gene expression. The degradation of BBX4 in darkness is dependent on CONSTITUTIVELY PHOTOMORPHOGENIC 1 (COP1) and the 26S proteasome system. Collectively, BBX4 acts as a key component of the phyB-PIF3-mediated signaling module and fine tunes the red light action. phyB promotes the accumulation of BBX4, which in turn serves to repress PIF3 action through direct physical interaction to promote photomorphogenic development in red light.
Keywords: BBX4; COP1; light signaling; photomorphogenesis; phyB.
Conflict of interest statement
The authors declare no competing interest.
Figures


References
-
- Sharrock R. A., Quail P. H., Novel phytochrome sequences in Arabidopsis thaliana: Structure, evolution, and differential expression of a plant regulatory photoreceptor family. Genes Dev. 3, 1745–1757 (1989). - PubMed
-
- Lin C., et al. , Association of flavin adenine dinucleotide with the Arabidopsis blue light receptor CRY1. Science 269, 968–970 (1995). - PubMed
-
- Guo H., Yang H., Mockler T. C., Lin C., Regulation of flowering time by Arabidopsis photoreceptors. Science 279, 1360–1363 (1998). - PubMed
-
- Rizzini L., et al. , Perception of UV-B by the Arabidopsis UVR8 protein. Science 332, 103–106 (2011). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
