

Despite mutation acquisition in hematopoietic stem cells, JMML-propagating cells are not always restricted to this compartment
- PMID: 31776464
- PMCID: PMC7266742
- DOI: 10.1038/s41375-019-0662-y
Despite mutation acquisition in hematopoietic stem cells, JMML-propagating cells are not always restricted to this compartment
Erratum in
-
Correction: Despite mutation acquisition in hematopoietic stem cells, JMML-propagating cells are not always restricted to this compartment.Leukemia. 2020 Jul;34(7):1973. doi: 10.1038/s41375-020-0720-5. Leukemia. 2020. PMID: 32005921 Free PMC article.
Abstract
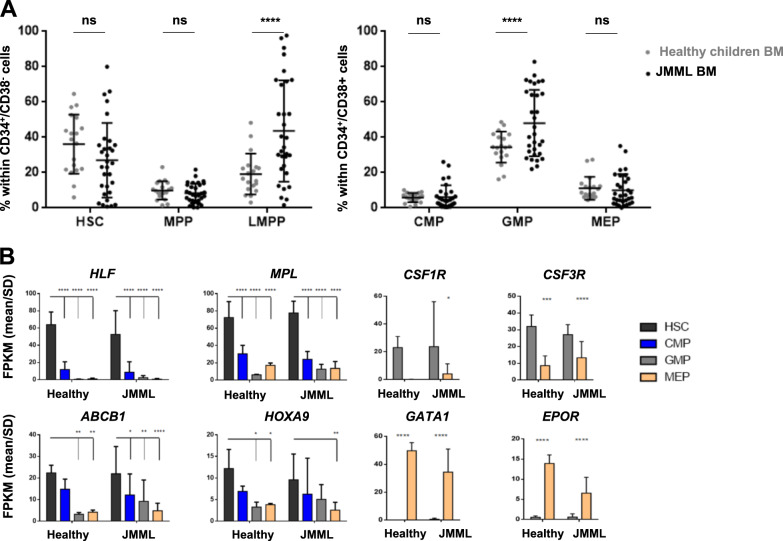
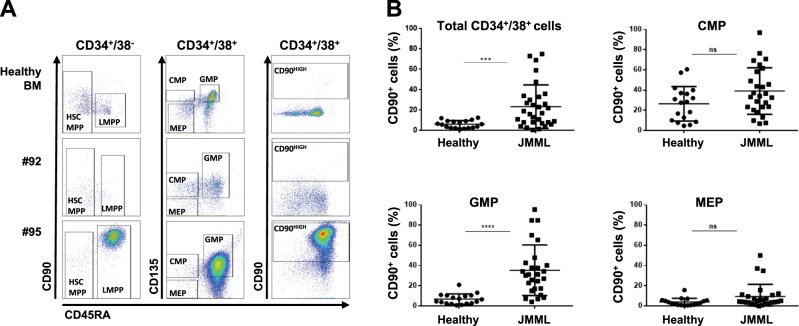
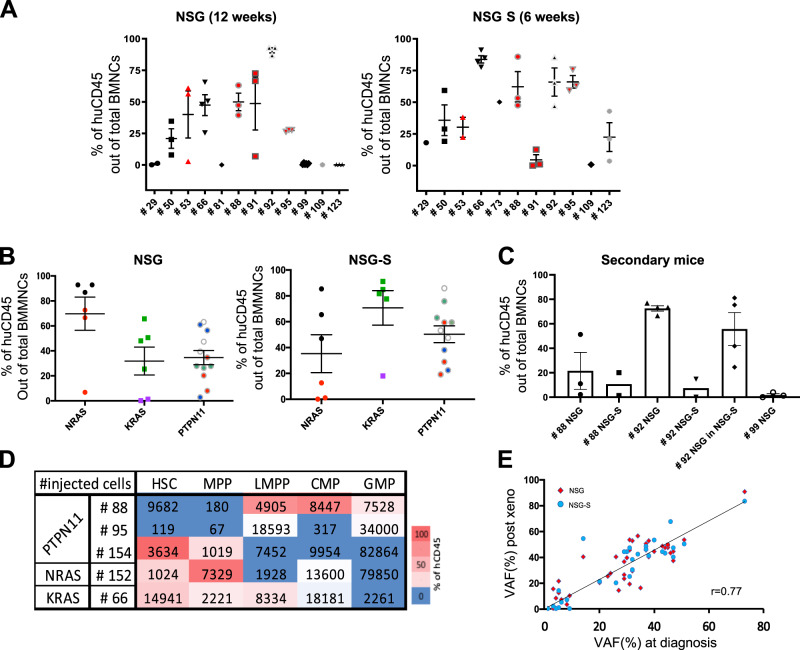
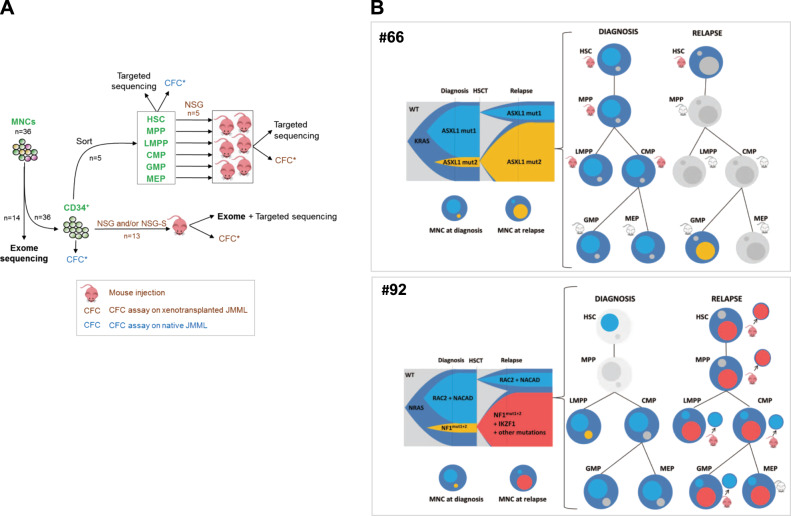
Juvenile myelomonocytic leukemia (JMML) is a rare aggressive myelodysplastic/myeloproliferative neoplasm of early childhood, initiated by RAS-activating mutations. Genomic analyses have recently described JMML mutational landscape; however, the nature of JMML-propagating cells (JMML-PCs) and the clonal architecture of the disease remained until now elusive. Combining genomic (exome, RNA-seq), Colony forming assay and xenograft studies, we detect the presence of JMML-PCs that faithfully reproduce JMML features including the complex/nonlinear organization of dominant/minor clones, both at diagnosis and relapse. Further integrated analysis also reveals that although the mutations are acquired in hematopoietic stem cells, JMML-PCs are not always restricted to this compartment, highlighting the heterogeneity of the disease during the initiation steps. We show that the hematopoietic stem/progenitor cell phenotype is globally maintained in JMML despite overexpression of CD90/THY-1 in a subset of patients. This study shed new lights into the ontogeny of JMML, and the identity of JMML-PCs, and provides robust models to monitor the disease and test novel therapeutic approaches.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
