

Monocarboxylate transporter 1 promotes classical microglial activation and pro-inflammatory effect via 6-phosphofructo-2-kinase/fructose-2, 6-biphosphatase 3
- PMID: 31779643
- PMCID: PMC6883695
- DOI: 10.1186/s12974-019-1648-4
Monocarboxylate transporter 1 promotes classical microglial activation and pro-inflammatory effect via 6-phosphofructo-2-kinase/fructose-2, 6-biphosphatase 3
Abstract
Background: Microglia, the resident macrophages of central nervous system, have been initially categorized into two opposite phenotypes: classical activation related to pro-inflammatory responses and alternative activation corresponding with anti-inflammatory reactions and tissue remodeling. The correlation between metabolic pattern and microglial activation has been identified. However, little is known about the mechanism of metabolism-mediated microglia polarization and pro-inflammatory effect.
Methods: Metabolic alteration was analyzed in different phenotypes of microglia in vitro. LPS-induced neuroinflammation and sickness behavior mouse model was used to investigate the effect of lactate on classical microglial activation in vivo.
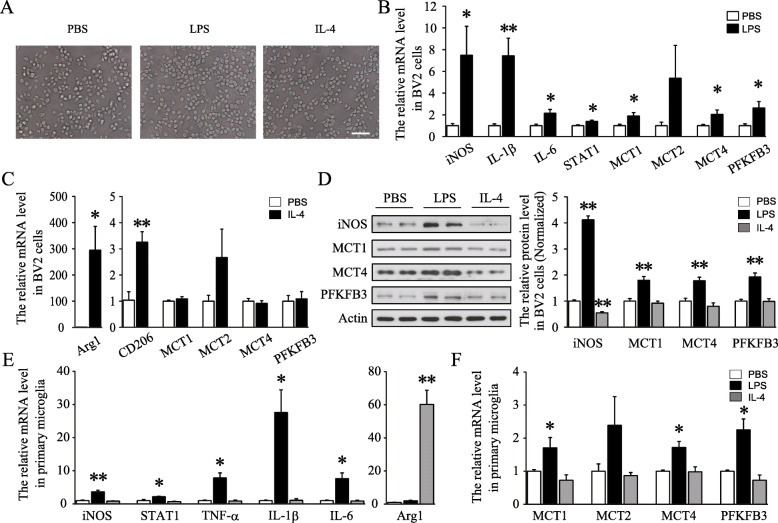
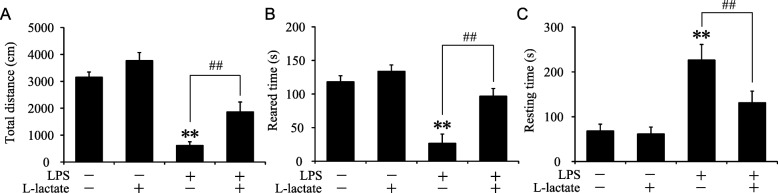
Results: Glycolysis-related regulators, monocarboxylate transporter 1 (MCT1), MCT4, and pro-glycolytic enzyme 6-phosphofructo-2-kinase/fructose-2, 6-biphosphatase 3 (PFKFB3), were specifically increased in LPS-stimulated primary microglia and microglia cell line BV2. Knockdown of MCT1 suppressed glycolysis rate and decreased LPS-induced expression of iNOS, interleukin-1β (IL-1β), IL-6, and phosphorylation of STAT1 in BV2 cells. Importantly, MCT1 promoted PFKFB3 expression via hypoxia-inducible factor-1α (Hif-1α), and overexpression of PFKFB3 restored the classical activation of BV2 cells suppressed by MCT1 silence. All above strongly suggested that MCT1/PFKFB3 might accelerate LPS-induced classical polarization of microglia probably by promoting glycolysis. Interestingly, additional administration of moderate lactate, which may block the transport function of MCT1, decreased LPS-induced classical activation and expression of PFKFB3 in BV2 cells. Intracerebroventricular injection of lactate ameliorated LPS-induced sickness behavior and classical polarization of microglia in mice.
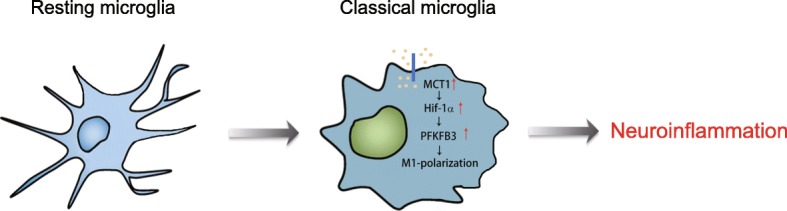
Conclusions: Our results demonstrate the key role of MCT1 in microglial classical activation and neuroinflammation in pathological conditions. In addition, lactate administration may be a potential therapy to suppress neuroinflammation by altering microglial polarization.
Keywords: Classical microglial polarization; Glycolysis; Lactate; MCT1; Neuroinflammation; PFKFB3.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
