

547 transcriptomes from 44 brain areas reveal features of the aging brain in non-human primates
- PMID: 31779658
- PMCID: PMC6883628
- DOI: 10.1186/s13059-019-1866-1
547 transcriptomes from 44 brain areas reveal features of the aging brain in non-human primates
Erratum in
-
Author Correction: 547 transcriptomes from 44 brain areas reveal features of the aging brain in non-human primates.Genome Biol. 2020 Feb 24;21(1):46. doi: 10.1186/s13059-020-01962-z. Genome Biol. 2020. PMID: 32093779 Free PMC article.
Abstract
Background: Brain aging is a complex process that depends on the precise regulation of multiple brain regions; however, the underlying molecular mechanisms behind this process remain to be clarified in non-human primates.
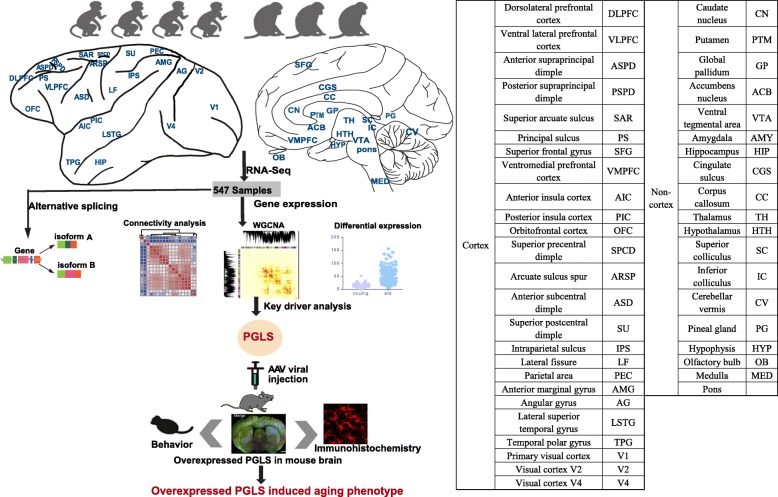
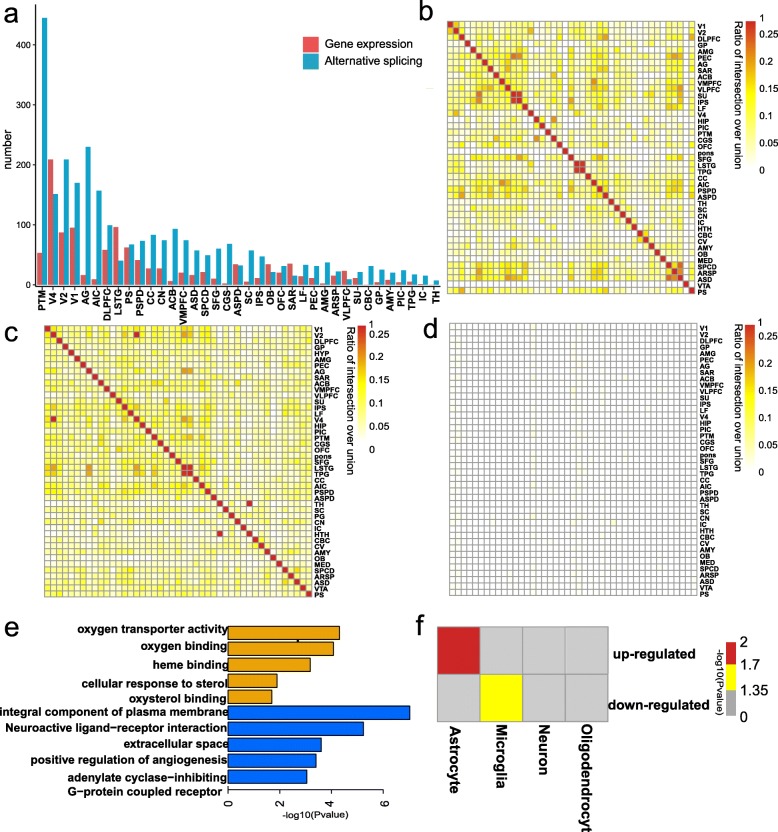
Results: Here, we explore non-human primate brain aging using 547 transcriptomes originating from 44 brain areas in rhesus macaques (Macaca mulatta). We show that expression connectivity between pairs of cerebral cortex areas as well as expression symmetry between the left and right hemispheres both decrease after aging. Although the aging mechanisms across different brain areas are largely convergent, changes in gene expression and alternative splicing vary at diverse genes, reinforcing the complex multifactorial basis of aging. Through gene co-expression network analysis, we identify nine modules that exhibit gain of connectivity in the aged brain and uncovered a hub gene, PGLS, underlying brain aging. We further confirm the functional significance of PGLS in mice at the gene transcription, molecular, and behavioral levels.
Conclusions: Taken together, our study provides comprehensive transcriptomes on multiple brain regions in non-human primates and provides novel insights into the molecular mechanism of healthy brain aging.
Keywords: Brain aging; Multiple brain regions; PGLS; Rhesus macaques; Transcriptome.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures




Similar articles
-
Regional and aging-specific cellular architecture of non-human primate brains.Genome Med. 2025 Apr 28;17(1):41. doi: 10.1186/s13073-025-01469-x. Genome Med. 2025. PMID: 40296047 Free PMC article.
-
A comprehensive transcriptional map of primate brain development.Nature. 2016 Jul 21;535(7612):367-75. doi: 10.1038/nature18637. Epub 2016 Jul 13. Nature. 2016. PMID: 27409810 Free PMC article.
-
The testicular transcriptome associated with spermatogonia differentiation initiated by gonadotrophin stimulation in the juvenile rhesus monkey (Macaca mulatta).Hum Reprod. 2017 Oct 1;32(10):2088-2100. doi: 10.1093/humrep/dex270. Hum Reprod. 2017. PMID: 28938749 Free PMC article.
-
Calorie restriction in nonhuman primates: assessing effects on brain and behavioral aging.Neuroscience. 2007 Apr 14;145(4):1359-64. doi: 10.1016/j.neuroscience.2006.10.031. Epub 2007 Jan 16. Neuroscience. 2007. PMID: 17223278 Review.
-
Aging and caloric restriction in nonhuman primates: behavioral and in vivo brain imaging studies.Ann N Y Acad Sci. 2001 Apr;928:316-26. doi: 10.1111/j.1749-6632.2001.tb05661.x. Ann N Y Acad Sci. 2001. PMID: 11795523 Review.
Cited by
-
Regional and aging-specific cellular architecture of non-human primate brains.Genome Med. 2025 Apr 28;17(1):41. doi: 10.1186/s13073-025-01469-x. Genome Med. 2025. PMID: 40296047 Free PMC article.
-
Multi-Omics Analysis Reveals Age-Related Microbial and Metabolite Alterations in Non-Human Primates.Microorganisms. 2023 Sep 26;11(10):2406. doi: 10.3390/microorganisms11102406. Microorganisms. 2023. PMID: 37894064 Free PMC article.
-
Gene co-expression analyses of health(span) across multiple species.NAR Genom Bioinform. 2022 Nov 29;4(4):lqac083. doi: 10.1093/nargab/lqac083. eCollection 2022 Dec. NAR Genom Bioinform. 2022. PMID: 36458022 Free PMC article.
-
SIRT2 counteracts primate cardiac aging via deacetylation of STAT3 that silences CDKN2B.Nat Aging. 2023 Oct;3(10):1269-1287. doi: 10.1038/s43587-023-00486-y. Epub 2023 Oct 2. Nat Aging. 2023. PMID: 37783815
-
Deficits in motor and cognitive functions in an adult mouse model of hypoxia-ischemia induced stroke.Sci Rep. 2020 Nov 26;10(1):20646. doi: 10.1038/s41598-020-77678-8. Sci Rep. 2020. PMID: 33244072 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
