

Mitochondria, Telomeres and Telomerase Subunits
- PMID: 31781563
- PMCID: PMC6851022
- DOI: 10.3389/fcell.2019.00274
Mitochondria, Telomeres and Telomerase Subunits
Abstract
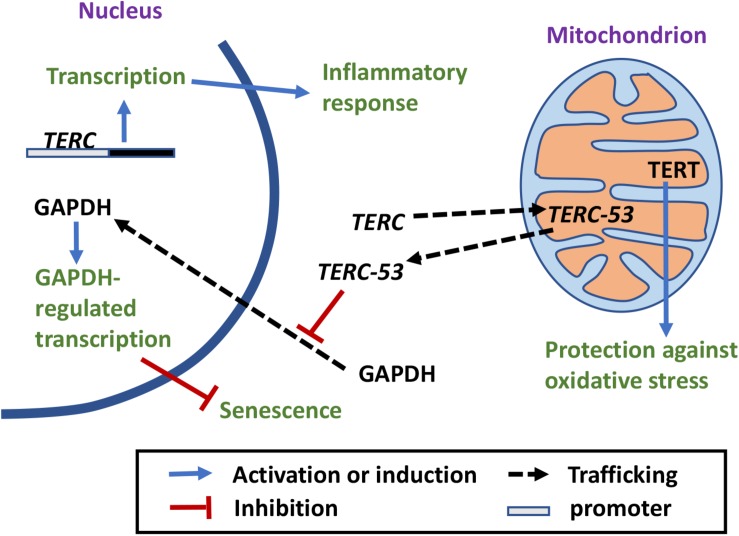
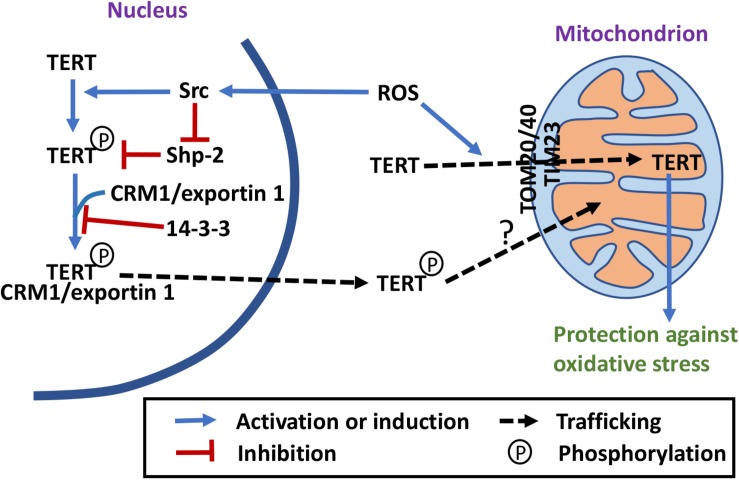
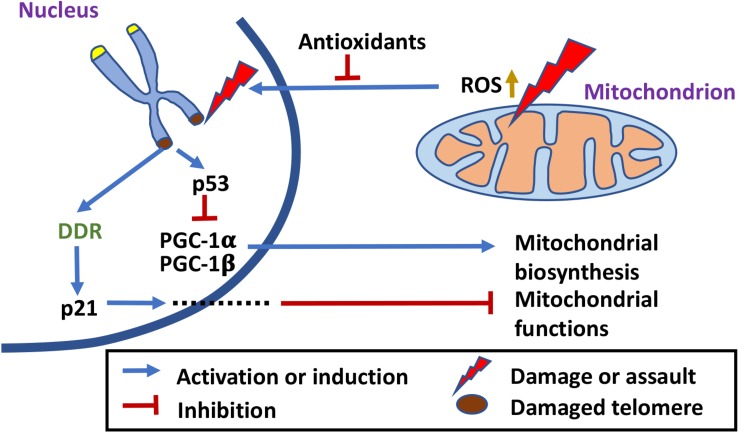
Mitochondrial functions and telomere functions have mostly been studied independently. In recent years, it, however, has become clear that there are intimate links between mitochondria, telomeres, and telomerase subunits. Mitochondrial dysfunctions cause telomere attrition, while telomere damage leads to reprogramming of mitochondrial biosynthesis and mitochondrial dysfunctions, which has important implications in aging and diseases. In addition, evidence has accumulated that telomere-independent functions of telomerase also exist and that the protein component of telomerase TERT shuttles between the nucleus and mitochondria under oxidative stress. Our previously published data show that the RNA component of telomerase TERC is also imported into mitochondria, processed, and exported back to the cytosol. These data show a complex regulation network where telomeres, nuclear genome, and mitochondria are co-regulated by multi-localization and multi-function proteins and RNAs. This review summarizes the connections between mitochondria and telomeres, the mitochondrion-related functions of telomerase subunits, and how they play a role in crosstalk between mitochondria and the nucleus.
Keywords: TERC; TERT; aging; mitochondria; telomerase; telomere.
Copyright © 2019 Zheng, Huang and Wang.
Figures
References
-
- Beckman J. S., Koppenol W. H. (1996). Nitric oxide, superoxide, and peroxynitrite: the good, the bad, and ugly. Am. J. Physiol. 271 C1424–C1437. - PubMed
Publication types
LinkOut - more resources
Full Text Sources