

Small RNA Mcr11 requires the transcription factor AbmR for stable expression and regulates genes involved in the central metabolism of Mycobacterium tuberculosis
- PMID: 31782837
- PMCID: PMC7064933
- DOI: 10.1111/mmi.14436
Small RNA Mcr11 requires the transcription factor AbmR for stable expression and regulates genes involved in the central metabolism of Mycobacterium tuberculosis
Abstract
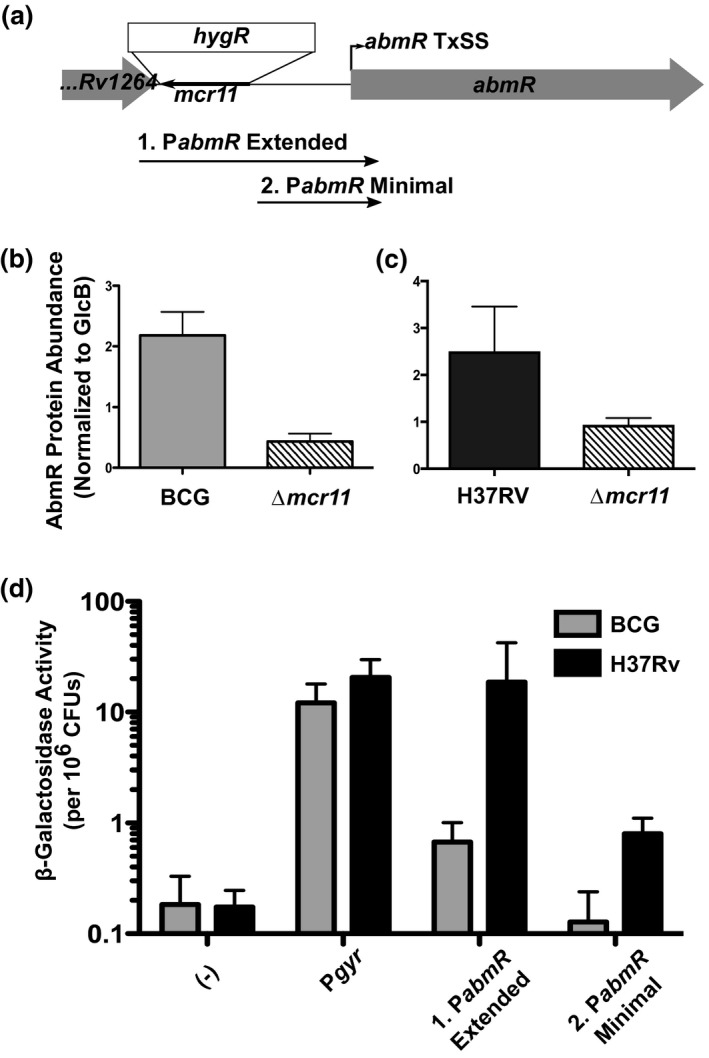
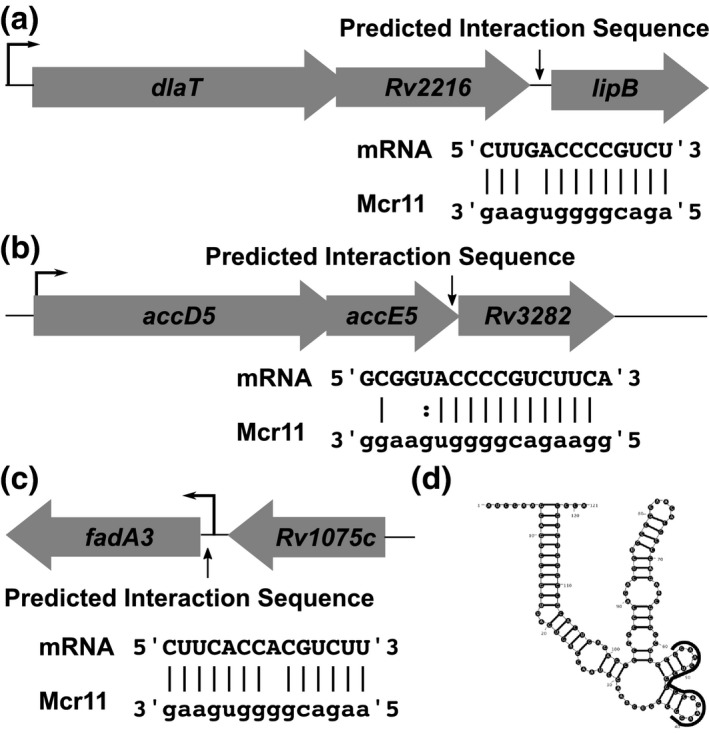
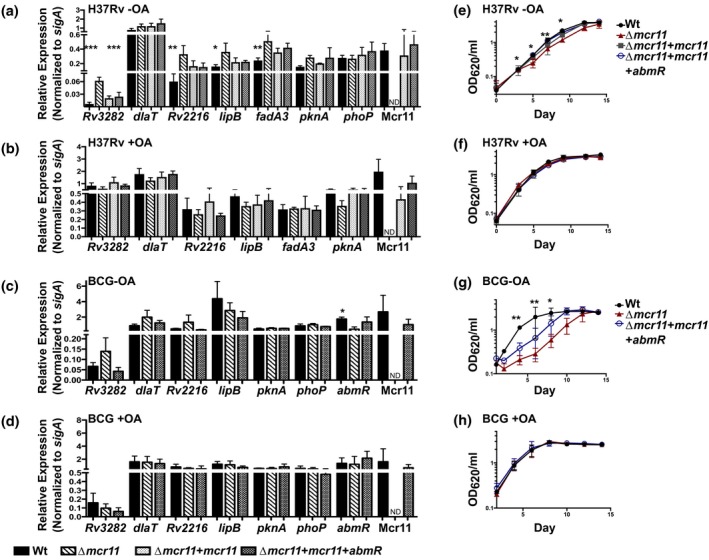
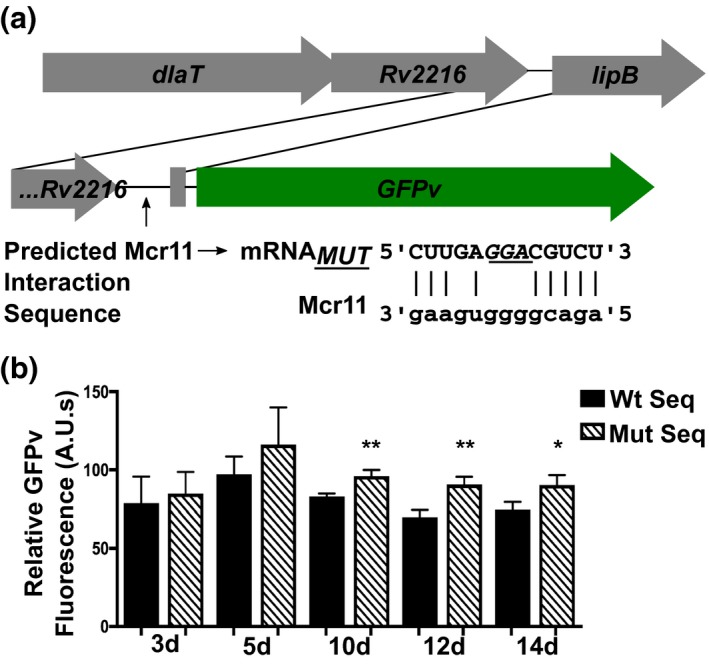
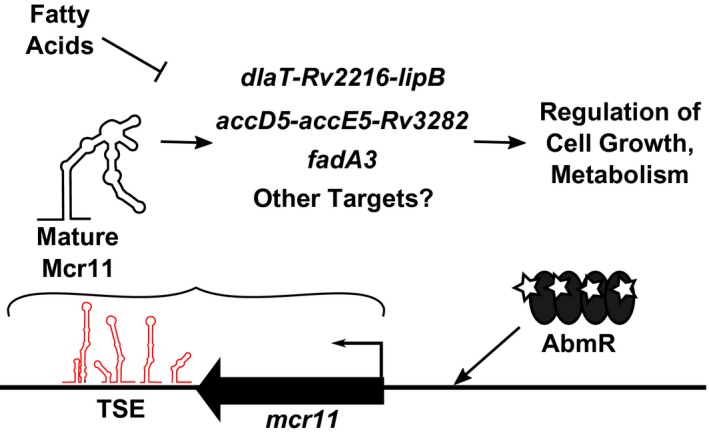
Mycobacterium tuberculosis (Mtb), the etiologic agent of tuberculosis, must adapt to host-associated environments during infection by modulating gene expression. Small regulatory RNAs (sRNAs) are key regulators of bacterial gene expression, but their roles in Mtb are not well understood. Here, we address the expression and function of the Mtb sRNA Mcr11, which is associated with slow bacterial growth and chronic infections in mice. We found that stable expression of Mcr11 requires multiple factors specific to TB-complex bacteria, including the AbmR transcription factor. Bioinformatic analyses used to predict regulatory targets of Mcr11 identified 7-11 nucleotide regions with potential for direct base-pairing with Mcr11 immediately upstream of Rv3282, fadA3, and lipB. mcr11-dependent regulation of these genes was demonstrated using qRT-PCR and found to be responsive to the presence of fatty acids. Mutation of the putative Mcr11 base-pairing site upstream of lipB in a promoter reporter strain resulted in significant de-repression of lipB expression, similar to that observed in mcr11-deleted Mtb. These studies establish Mcr11's roles in regulating growth and central metabolism in Mtb. Our finding that multiple TB-complex-specific factors are required for production of stable Mcr11 also emphasizes the need to better understand mechanisms of sRNA expression and stability in TB.
Keywords: RNA stability; RNA termination; gene regulation; lipoylation; sRNA targets.
© 2019 The Authors. Molecular Microbiology published by John Wiley & Sons Ltd.
Conflict of interest statement
None declared.
Figures
References
-
- Abendroth, J. , Ollodart, A. , Andrews, E. S. , Myler, P. J. , Staker, B. L. , Edwards, T. E. , … Grundner, C. (2014). Mycobacterium tuberculosis Rv2179c protein establishes a new exoribonuclease family with broad phylogenetic distribution. Journal of Biological Chemistry, 289, 2139–2147. - PMC - PubMed
-
- Arnvig, K. B. , Comas, I. , Thomson, N. R. , Houghton, J. , Boshoff, H. I. , Croucher, N. J. , … Young, D. B. (2011). Sequence‐based analysis uncovers an abundance of non‐coding RNA in the total transcriptome of Mycobacterium tuberculosis . PLoS Path, 7, e1002342 10.1371/journal.ppat.1002342 - DOI - PMC - PubMed
-
- Bardarov, S. , Bardarov, S., Jr. , Pavelka, M. S., Jr., Sambandamurthy, V. , Larsen, M. , Tufariello, J. , … Jacobs, W. R., Jr. (2002). Specialized transduction: An efficient method for generating marked and unmarked targeted gene disruptions in Mycobacterium tuberculosis, M. bovis BCG and M. smegmatis. Microbiology, 148, 3007–3017. 10.1099/00221287-148-10-3007 - DOI - PubMed
-
- Bazet Lyonnet, B. , Diacovich, L. , Cabruja, M. , Bardou, F. , Quemard, A. , Gago, G. , & Gramajo, H. (2014). Pleiotropic effect of AccD5 and AccE5 depletion in acyl‐coenzyme A carboxylase activity and in lipid biosynthesis in mycobacteria. PLoS ONE, 9, e99853 10.1371/journal.pone.0099853 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
