

Toxin-Triggered Interleukin-1 Receptor Signaling Enables Early-Life Discrimination of Pathogenic versus Commensal Skin Bacteria
- PMID: 31784259
- PMCID: PMC6989301
- DOI: 10.1016/j.chom.2019.10.007
Toxin-Triggered Interleukin-1 Receptor Signaling Enables Early-Life Discrimination of Pathogenic versus Commensal Skin Bacteria
Abstract
The host must develop tolerance to commensal microbes and protective responses to infectious pathogens, yet the mechanisms enabling a privileged relationship with commensals remain largely unknown. Skin colonization by commensal Staphylococcus epidermidis facilitates immune tolerance preferentially in neonates via induction of antigen-specific regulatory T cells (Tregs). Here, we demonstrate that this tolerance is not indiscriminately extended to all bacteria encountered in this early window. Rather, neonatal colonization by Staphylococcus aureus minimally enriches for antigen-specific Tregs and does not prevent skin inflammation upon later-life exposure. S. aureus α-toxin contributes to this response by stimulating myeloid cell production of IL-1β, which limits S. aureus-specific Tregs. Loss of α-toxin or the IL-1 receptor increases Treg enrichment, whereas topical application of IL-1β or α-toxin diminishes tolerogenic responses to S. epidermidis. Thus, the preferential activation of a key alarmin pathway facilitates early discrimination of microbial "foe" from "friend," thereby preventing tolerance to a common skin pathogen.
Keywords: IL-1; commensal; dendritic cells; neonatal; pathogen; regulatory T cells; skin bacteria; skin immunity; staphylococcus.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
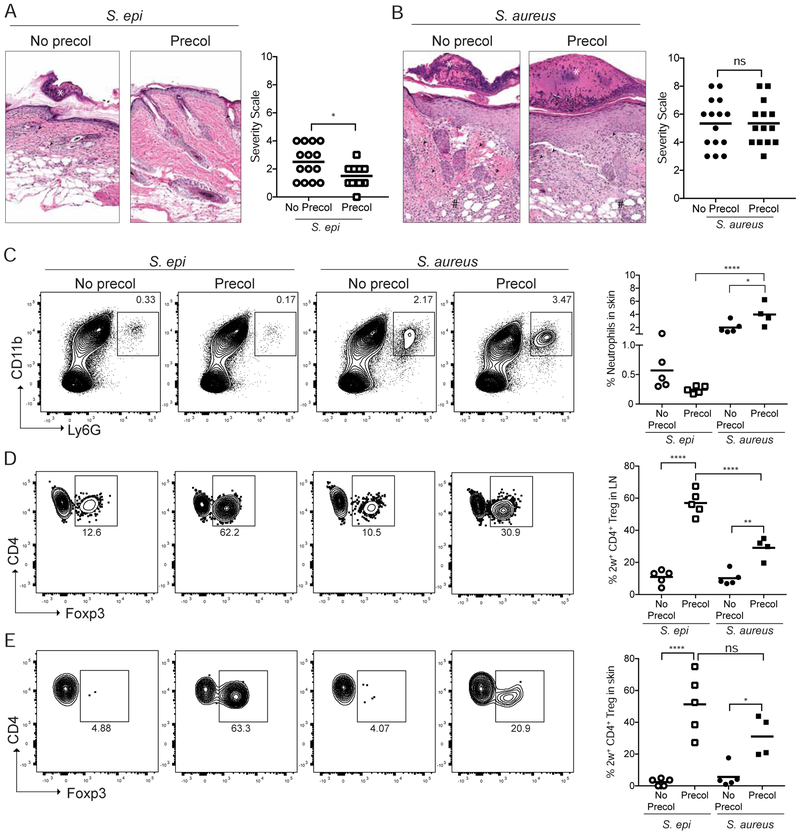
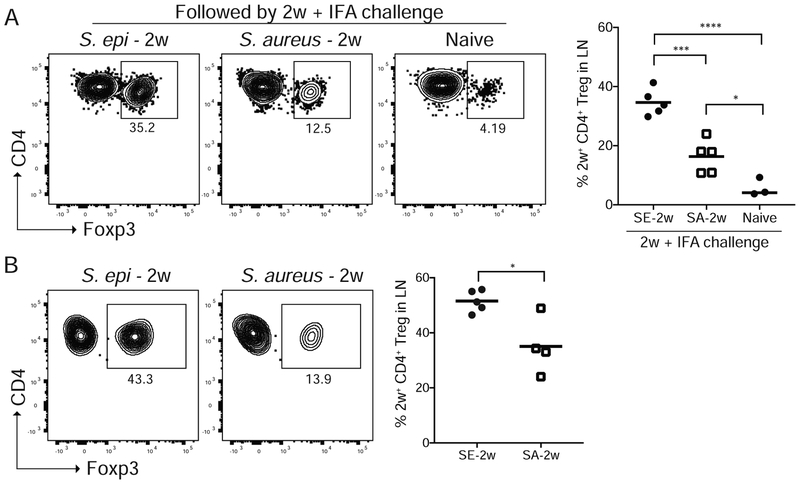
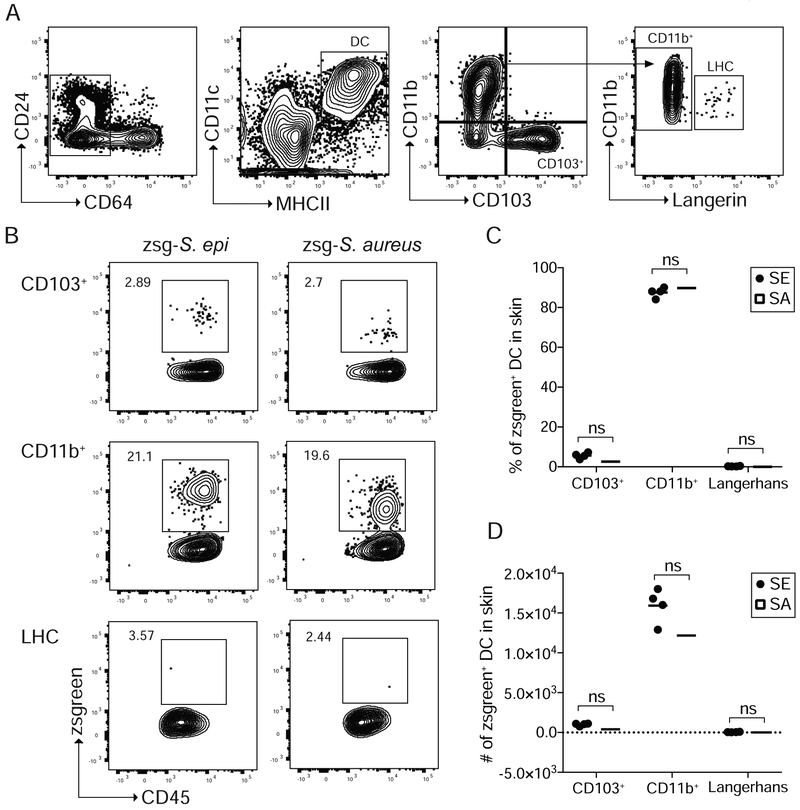
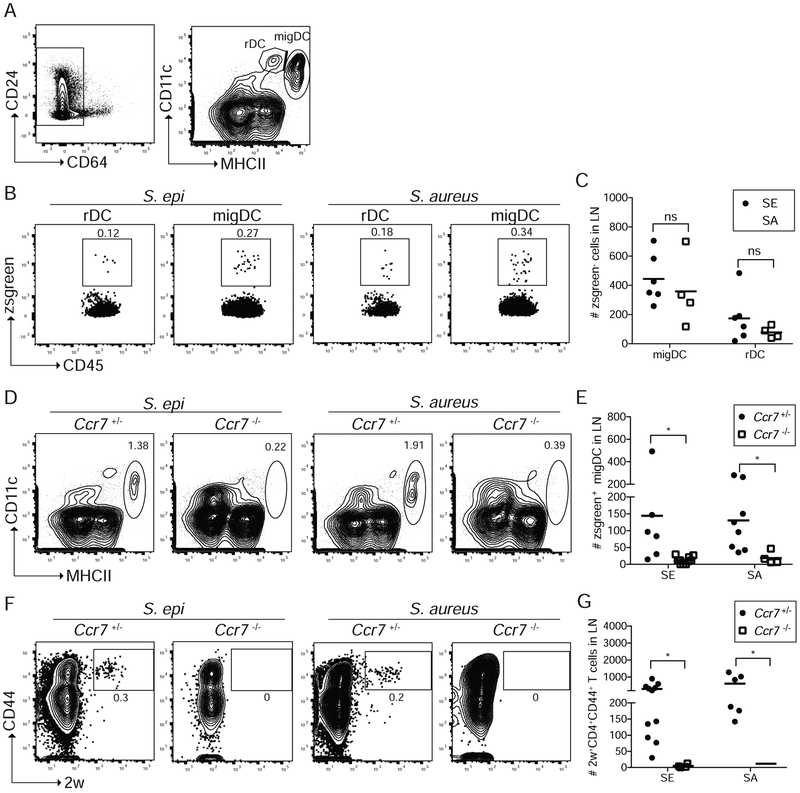

Comment in
-
Baby's First Bacteria: Discriminating Colonizing Commensals from Pathogens.Cell Host Microbe. 2019 Dec 11;26(6):705-707. doi: 10.1016/j.chom.2019.11.008. Cell Host Microbe. 2019. PMID: 31951581
Similar articles
-
Look Who's Talking: Host and Pathogen Drivers of Staphylococcus epidermidis Virulence in Neonatal Sepsis.Int J Mol Sci. 2022 Jan 13;23(2):860. doi: 10.3390/ijms23020860. Int J Mol Sci. 2022. PMID: 35055041 Free PMC article. Review.
-
A Wave of Regulatory T Cells into Neonatal Skin Mediates Tolerance to Commensal Microbes.Immunity. 2015 Nov 17;43(5):1011-21. doi: 10.1016/j.immuni.2015.10.016. Immunity. 2015. PMID: 26588783 Free PMC article.
-
Establishing Tolerance to Commensal Skin Bacteria: Timing Is Everything.Dermatol Clin. 2017 Jan;35(1):1-9. doi: 10.1016/j.det.2016.07.007. Dermatol Clin. 2017. PMID: 27890233 Free PMC article. Review.
-
Staphylococcus aureus Epicutaneous Exposure Drives Skin Inflammation via IL-36-Mediated T Cell Responses.Cell Host Microbe. 2017 Nov 8;22(5):653-666.e5. doi: 10.1016/j.chom.2017.10.006. Cell Host Microbe. 2017. PMID: 29120743 Free PMC article.
-
"Bringing Up Baby" to Tolerate Germs.Immunity. 2015 Nov 17;43(5):842-4. doi: 10.1016/j.immuni.2015.10.020. Immunity. 2015. PMID: 26588777
Cited by
-
Human Respiratory and Gut Microbiomes-Do They Really Contribute to Respiratory Health?Front Pediatr. 2020 Sep 3;8:528. doi: 10.3389/fped.2020.00528. eCollection 2020. Front Pediatr. 2020. PMID: 33014929 Free PMC article. Review.
-
Look Who's Talking: Host and Pathogen Drivers of Staphylococcus epidermidis Virulence in Neonatal Sepsis.Int J Mol Sci. 2022 Jan 13;23(2):860. doi: 10.3390/ijms23020860. Int J Mol Sci. 2022. PMID: 35055041 Free PMC article. Review.
-
Immunometabolism: new insights and lessons from antigen-directed cellular immune responses.Semin Immunopathol. 2020 Jun;42(3):279-313. doi: 10.1007/s00281-020-00798-w. Epub 2020 Jun 9. Semin Immunopathol. 2020. PMID: 32519148 Free PMC article. Review.
-
The Role of Coagulase-Negative Staphylococci Biofilms on Late-Onset Sepsis: Current Challenges and Emerging Diagnostics and Therapies.Antibiotics (Basel). 2023 Mar 10;12(3):554. doi: 10.3390/antibiotics12030554. Antibiotics (Basel). 2023. PMID: 36978421 Free PMC article. Review.
-
Staphylococcus aureus-specific TIGIT+ Treg are present in the blood of healthy subjects - a hurdle for vaccination?Front Immunol. 2025 Feb 4;15:1500696. doi: 10.3389/fimmu.2024.1500696. eCollection 2024. Front Immunol. 2025. PMID: 39981298 Free PMC article.
References
-
- Allgaier H, Jung G, Werner RG, Schneider U, Zähner H, 1986. Epidermin: sequencing of a heterodetic tetracyclic 21-peptide amide antibiotic. Eur. J. Biochem 160, 9–22. - PubMed
-
- Anandasabapathy N, Feder R, Mollah S, Tse S-W, Longhi MP, Mehandru S, Matos I, Cheong C, Ruane D, Brane L, Teixeira A, Dobrin J, Mizenina O, Park CG, Meredith M, Clausen BE, Nussenzweig MC, Steinman RM, 2014. Classical Flt3L-dependent dendritic cells control immunity to protein vaccine. J. Exp. Med 211, 1875–1891. doi:10.1084/jem.20131397 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
