

Branched Photoswitchable Tethered Ligands Enable Ultra-efficient Optical Control and Detection of G Protein-Coupled Receptors In Vivo
- PMID: 31784287
- PMCID: PMC7216301
- DOI: 10.1016/j.neuron.2019.10.036
Branched Photoswitchable Tethered Ligands Enable Ultra-efficient Optical Control and Detection of G Protein-Coupled Receptors In Vivo
Abstract
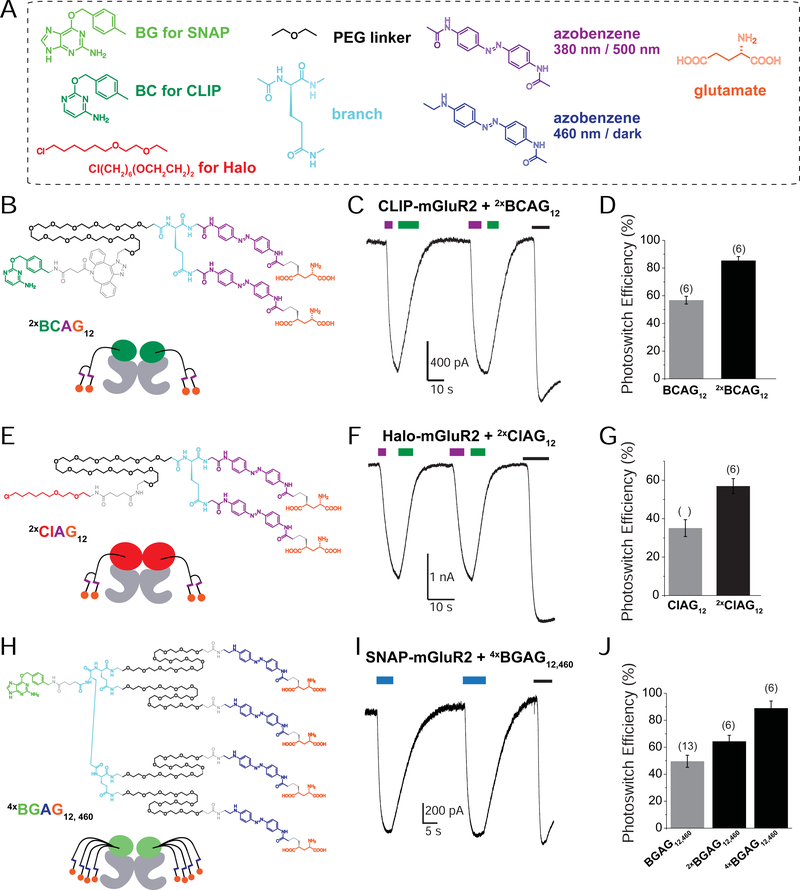
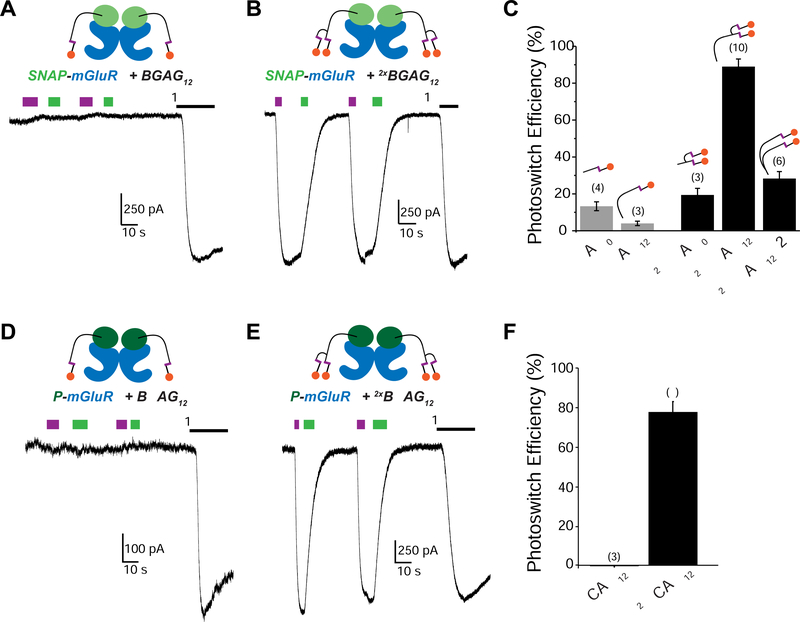
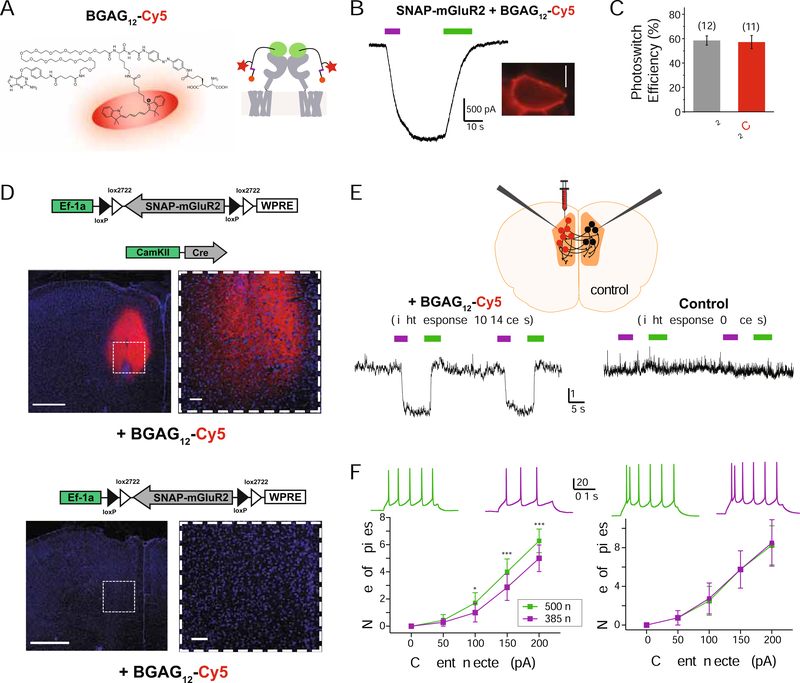
The limitations of classical drugs have spurred the development of covalently tethered photoswitchable ligands to control neuromodulatory receptors. However, a major shortcoming of tethered photopharmacology is the inability to obtain optical control with an efficacy comparable with that of the native ligand. To overcome this, we developed a family of branched photoswitchable compounds to target metabotropic glutamate receptors (mGluRs). These compounds permit photo-agonism of Gi/o-coupled group II mGluRs with near-complete efficiency relative to glutamate when attached to receptors via a range of orthogonal, multiplexable modalities. Through a chimeric approach, branched ligands also allow efficient optical control of Gq-coupled mGluR5, which we use to probe the spatiotemporal properties of receptor-induced calcium oscillations. In addition, we report branched, photoswitch-fluorophore compounds for simultaneous receptor imaging and manipulation. Finally, we demonstrate this approach in vivo in mice, where photoactivation of SNAP-mGluR2 in the medial prefrontal cortex reversibly modulates working memory in normal and disease-associated states.
Keywords: G protein-coupled receptor; astrocyte; calcium signaling; metabotropic glutamate receptor; neuromodulation; optogenetics; photopharmacology; prefrontal cortex; psychosis; working memory.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures





Comment in
-
Photocontrol of Metabotropic Glutamate Receptors: When One Agonist Is Not Enough, Make It Two.Neuron. 2020 Feb 5;105(3):395-397. doi: 10.1016/j.neuron.2020.01.013. Neuron. 2020. PMID: 32027826
References
-
- Aultman JM, and Moghaddam B. (2001). Distinct contributions of glutamate and dopamine receptors to temporal aspects of rodent working memory using a clinically relevant task. Psychopharmacology (Berl) 153, 353–364. - PubMed
-
- Bocchio M, Lukacs IP, Stacey R, Plaha P, Apostolopoulos V, Livermore L, Sen A, Ansorge O, Gillies MJ, Somogyi P, et al. (2018). Group II Metabotropic Glutamate Receptors Mediate Presynaptic Inhibition of Excitatory Transmission in Pyramidal Neurons of the Human Cerebral Cortex. Front Cell Neurosci 12, 508. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
