

Human iPSC-derived astrocytes from ALS patients with mutated C9ORF72 show increased oxidative stress and neurotoxicity
- PMID: 31787569
- PMCID: PMC6921360
- DOI: 10.1016/j.ebiom.2019.11.026
Human iPSC-derived astrocytes from ALS patients with mutated C9ORF72 show increased oxidative stress and neurotoxicity
Abstract
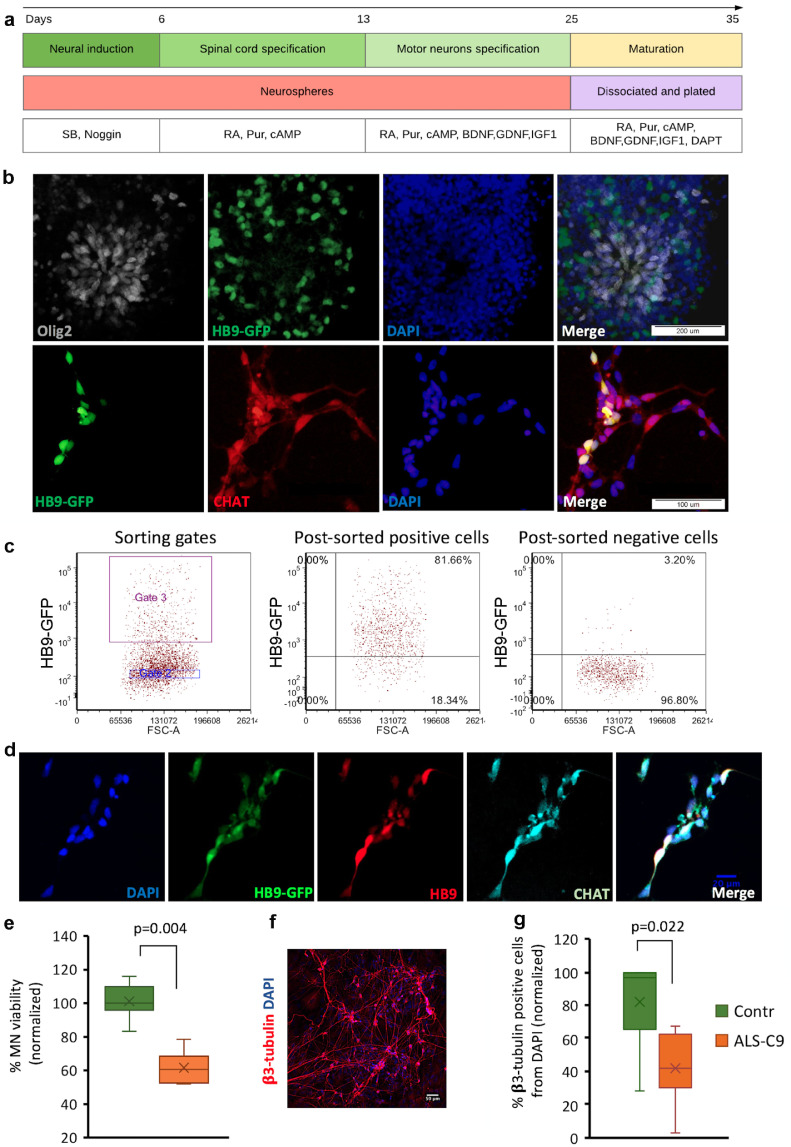
Background: Amyotrophic lateral sclerosis (ALS) is a progressive neurodegenerative disease that affects motor neurons (MNs). It was shown that human astrocytes with mutations in genes associated with ALS, like C9orf72 (C9) or SOD1, reduce survival of MNs. Astrocyte toxicity may be related to their dysfunction or the release of neurotoxic factors.
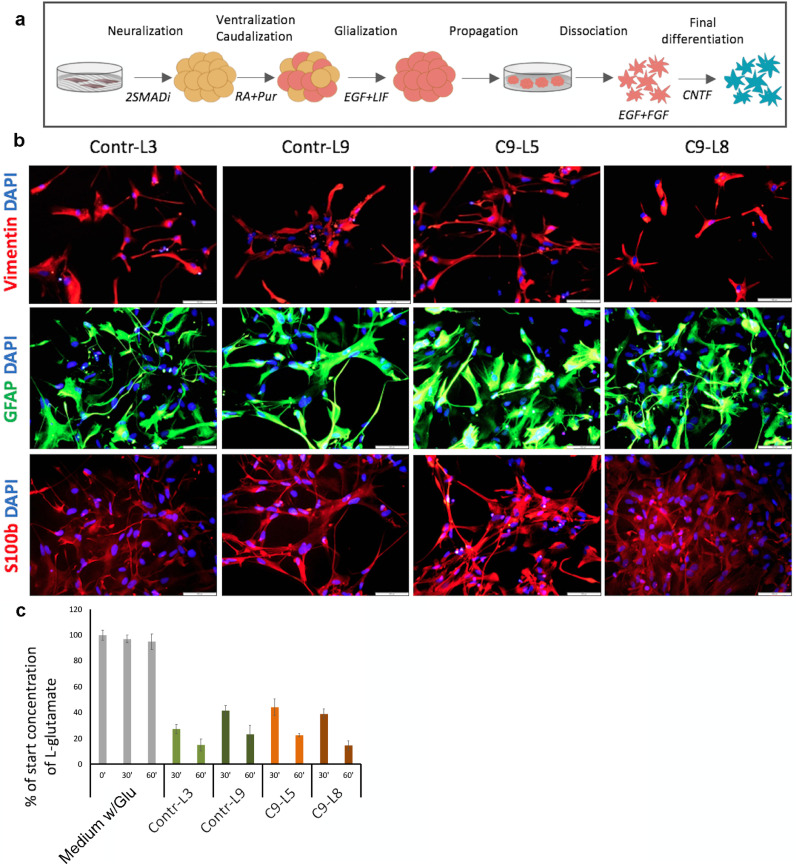
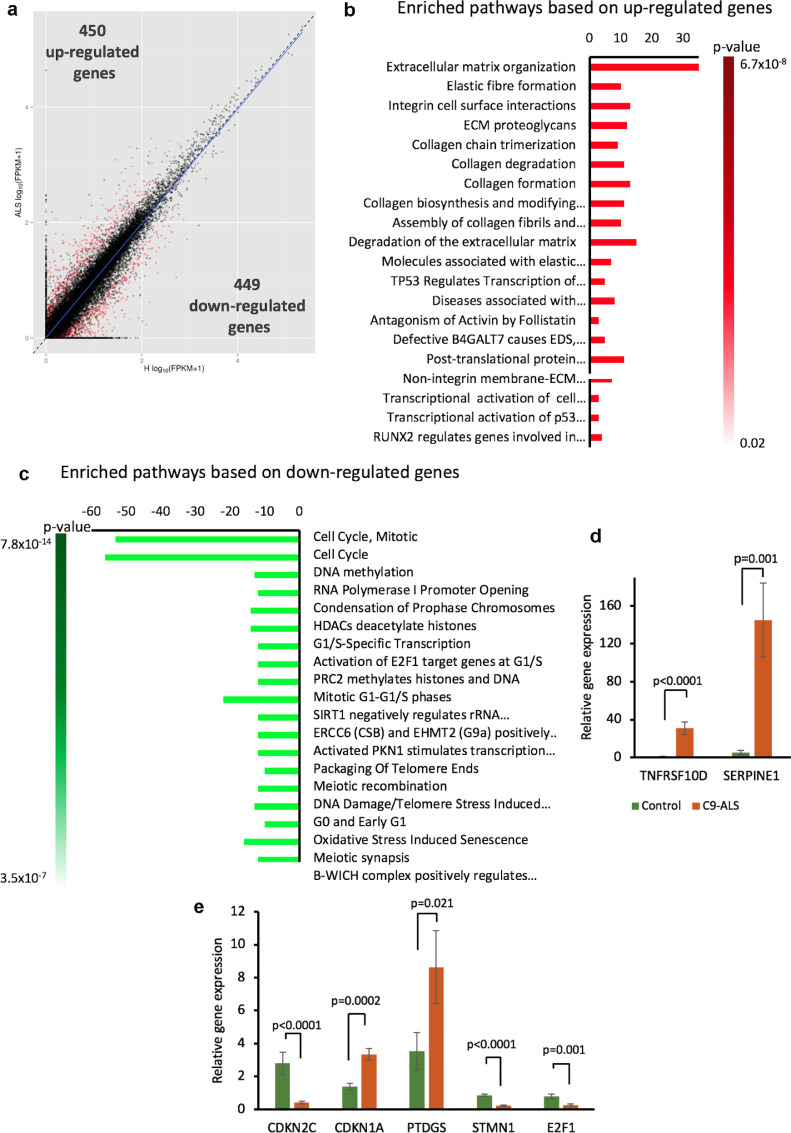
Methods: We used human induced pluripotent stem cell-derived astrocytes from ALS patients carrying C9orf72 mutations and non-affected donors. We utilized these cells to investigate astrocytic induced neuronal toxicity, changes in astrocyte transcription profile as well as changes in secretome profiles.
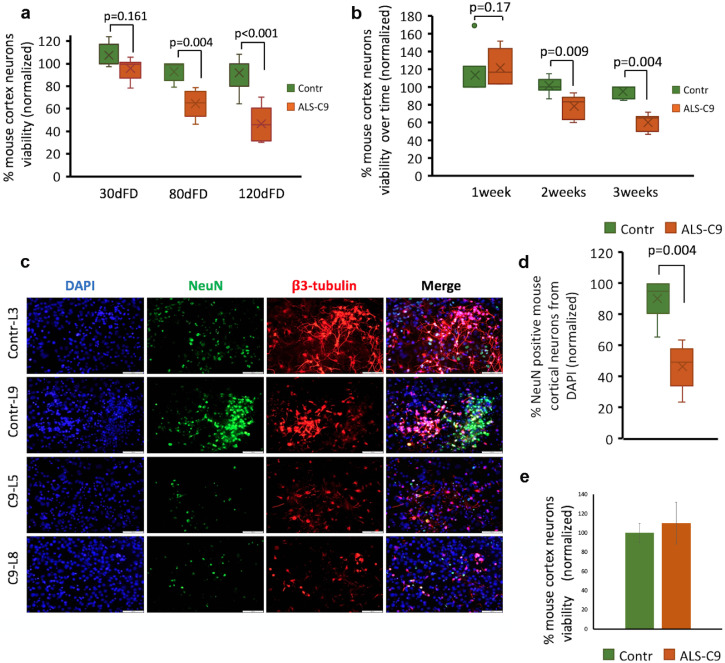
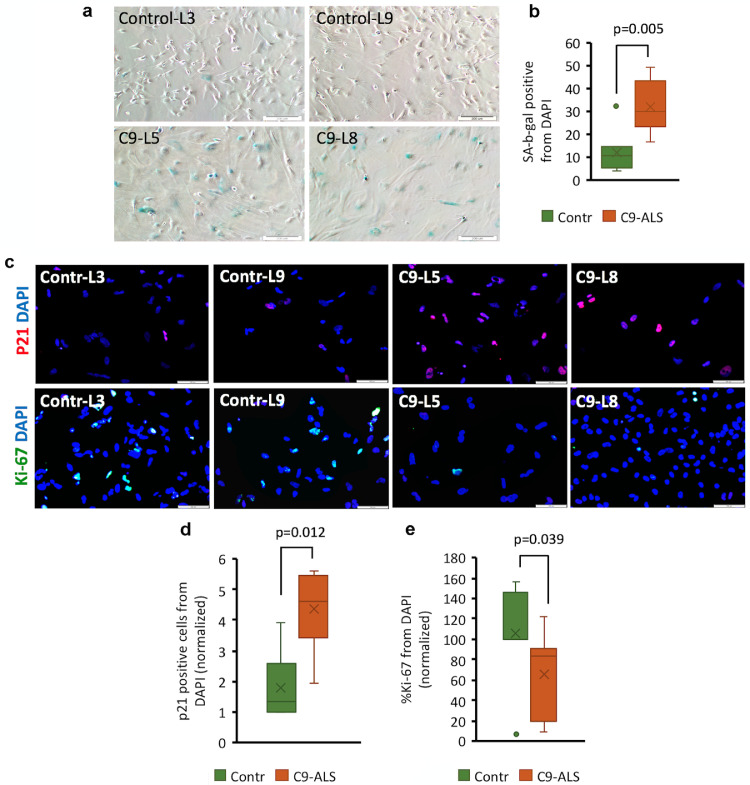
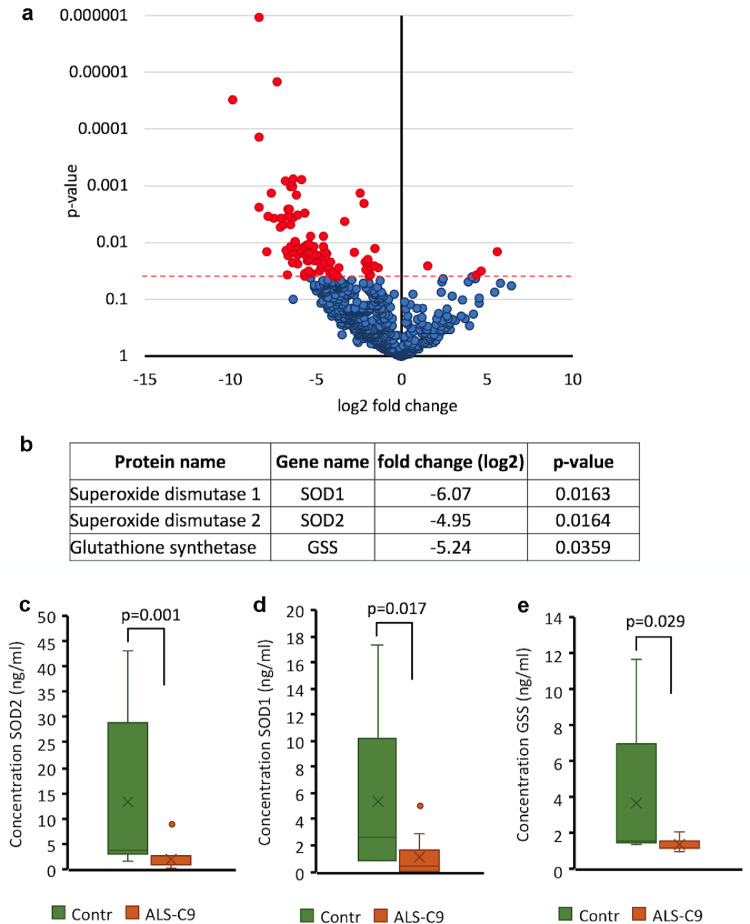
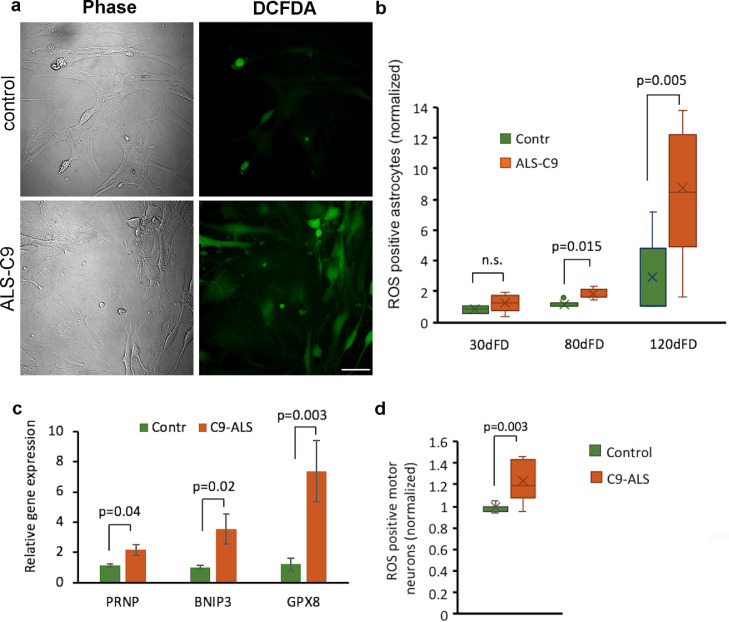
Findings: We report that C9-mutated astrocytes are toxic to MNs via soluble factors. The toxic effects of astrocytes are positively correlated with the length of astrocyte propagation in culture, consistent with the age-related nature of ALS. We show that C9-mutated astrocytes downregulate the secretion of several antioxidant proteins. In line with these findings, we show increased astrocytic oxidative stress and senescence. Importantly, media conditioned by C9-astrocytes increased oxidative stress in wild type MNs.
Interpretation: Our results suggest that dysfunction of C9-astrocytes leads to oxidative stress of themselves and MNs, which probably contributes to neurodegeneration. Our findings suggest that therapeutic strategies in familial ALS must not only target MNs but also focus on astrocytes to abrogate nervous system injury.
Keywords: Amyotrophic lateral sclerosis; Astrocytes; Neurotoxicity; Oxidative stress; Senescence; iPSC.
Copyright © 2019 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Benjamin Reubinoff is a member of the scientific advisory board of Kadimastem Ltd. and holds options in the company. A major focus of the company is the development of hESC-derived astrocytes for transplantation therapy in ALS. Michal Izrael is an employee of Kadimastem Ltd. and holds options in the company.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
