

The Calmodulin Binding Region of the Synaptic Vesicle Protein Mover Is Required for Homomeric Interaction and Presynaptic Targeting
- PMID: 31787876
- PMCID: PMC6856015
- DOI: 10.3389/fnmol.2019.00249
The Calmodulin Binding Region of the Synaptic Vesicle Protein Mover Is Required for Homomeric Interaction and Presynaptic Targeting
Abstract
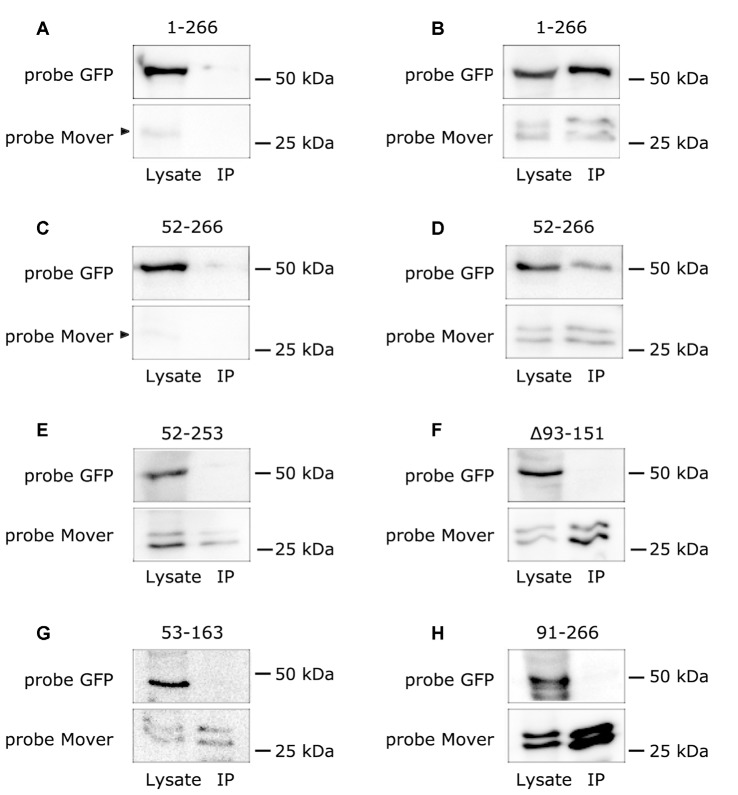
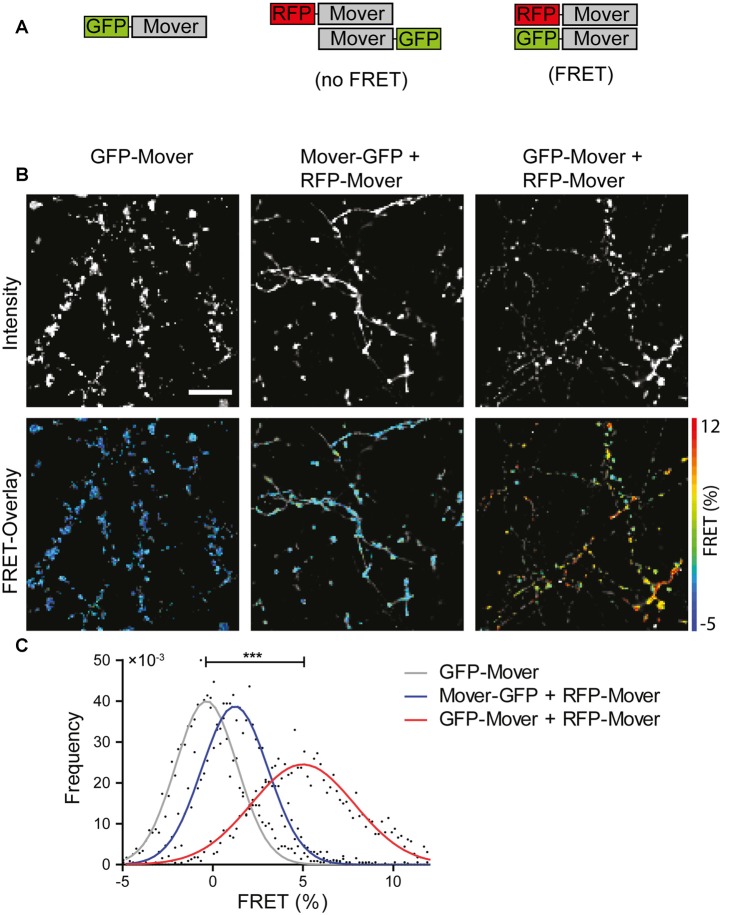
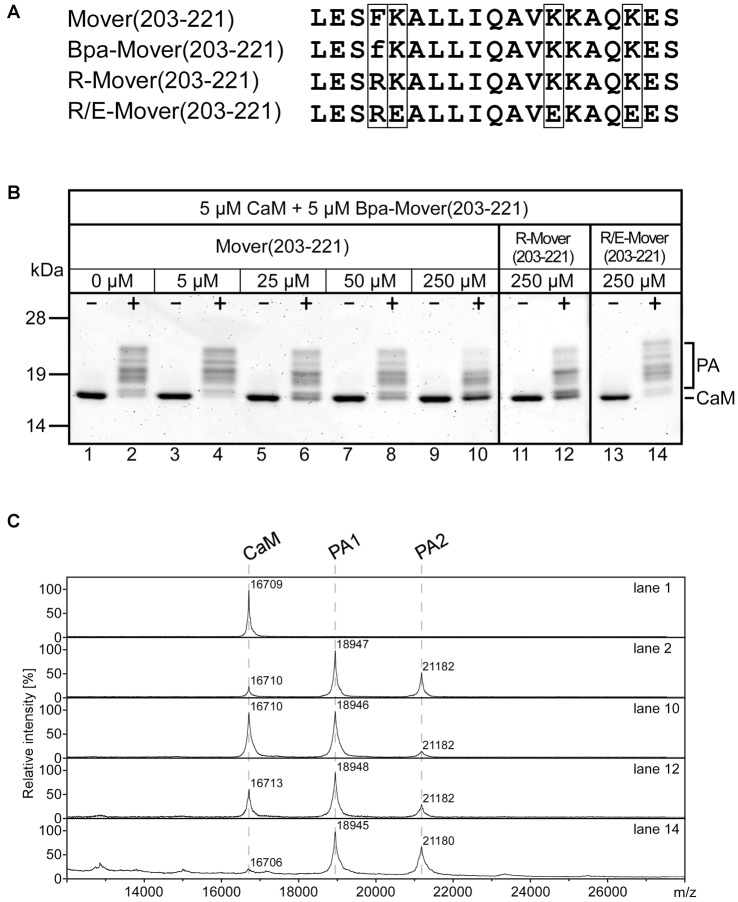
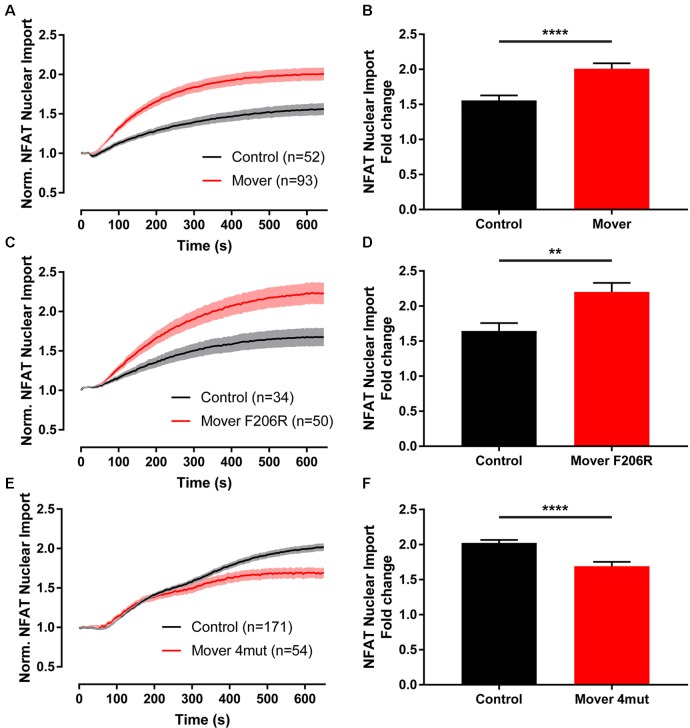
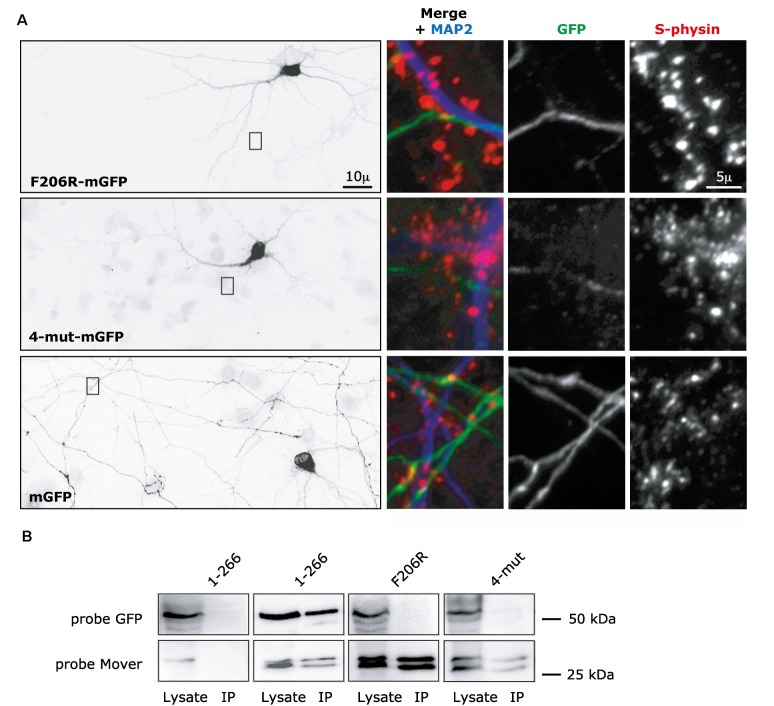
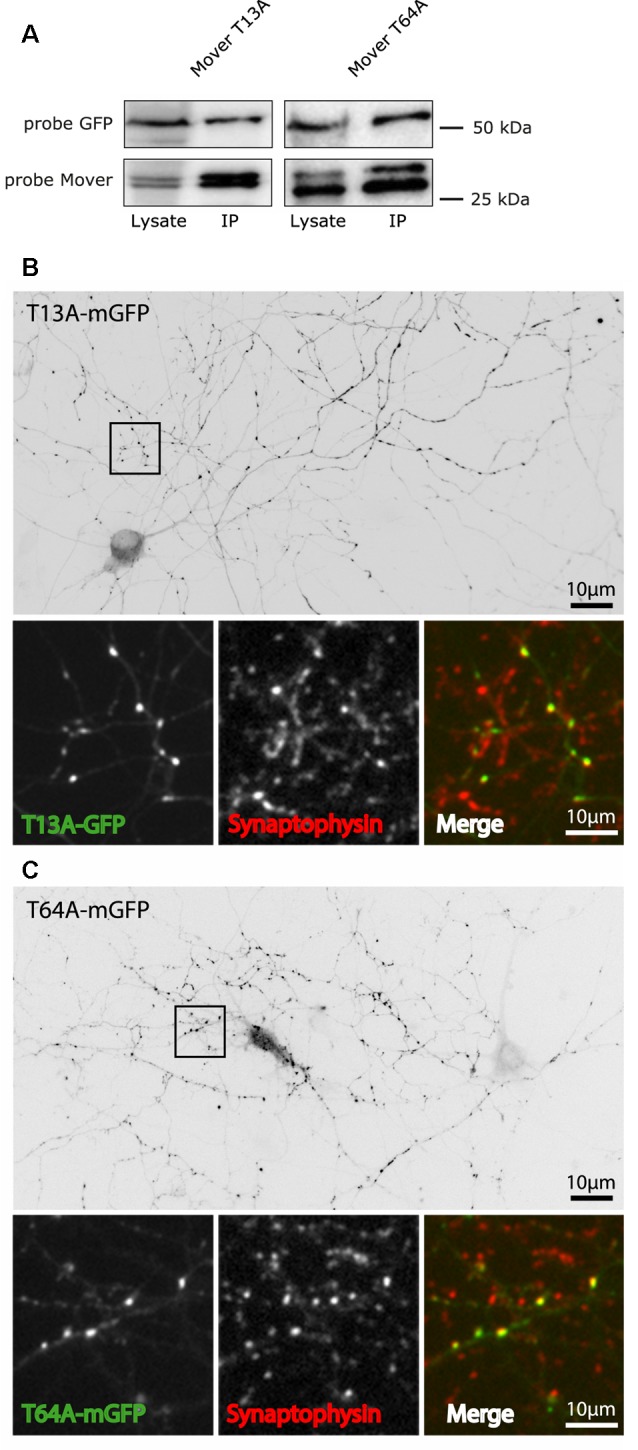
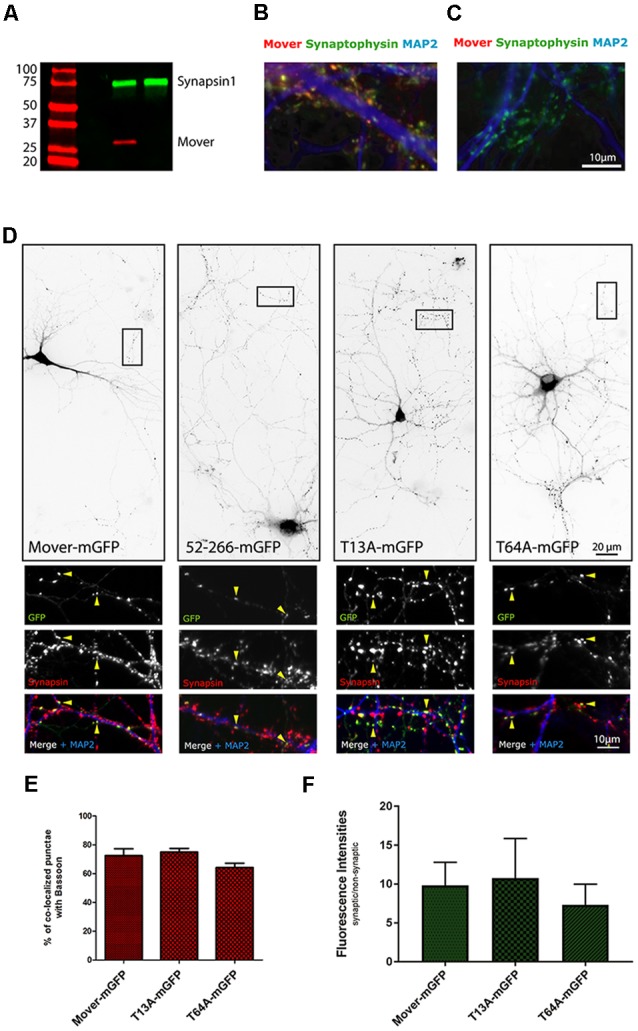
Neurotransmitter release is mediated by an evolutionarily conserved machinery. The synaptic vesicle (SV) associated protein Mover/TPRGL/SVAP30 does not occur in all species and all synapses. Little is known about its molecular properties and how it may interact with the conserved components of the presynaptic machinery. Here, we show by deletion analysis that regions required for homomeric interaction of Mover are distributed across the entire molecule, including N-terminal, central and C-terminal regions. The same regions are also required for the accumulation of Mover in presynaptic terminals of cultured neurons. Mutating two phosphorylation sites in N-terminal regions did not affect these properties. In contrast, a point mutation in the predicted Calmodulin (CaM) binding sequence of Mover abolished both homomeric interaction and presynaptic targeting. We show that this sequence indeed binds Calmodulin, and that recombinant Mover increases Calmodulin signaling upon heterologous expression. Our data suggest that presynaptic accumulation of Mover requires homomeric interaction mediated by regions distributed across large areas of the protein, and corroborate the hypothesis that Mover functionally interacts with Calmodulin signaling.
Keywords: TPRGL; calmodulin; mover; presynaptic; synaptic vesicle.
Copyright © 2019 Akula, Zhang, Viotti, Nestvogel, Rhee, Ebrecht, Reim, Wouters, Liepold, Jahn, Bogeski and Dresbach.
Figures

References
-
- Barber P. R., Ameee-Beg S. M., Gilbey J., Carlin L. M., Keppler M., Ng T. C., et al. (2009). Multiphoton time-domain fluorescence lifetime imaging microscopy: practical application to protein-protein interactions using global analysis. J. R. Soc. Interface 6, S93–S105. 10.1098/rsif.2008.0451.focus - DOI
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials