

Molecular structure in biomolecular condensates
- PMID: 31790873
- PMCID: PMC7117980
- DOI: 10.1016/j.sbi.2019.09.007
Molecular structure in biomolecular condensates
Abstract
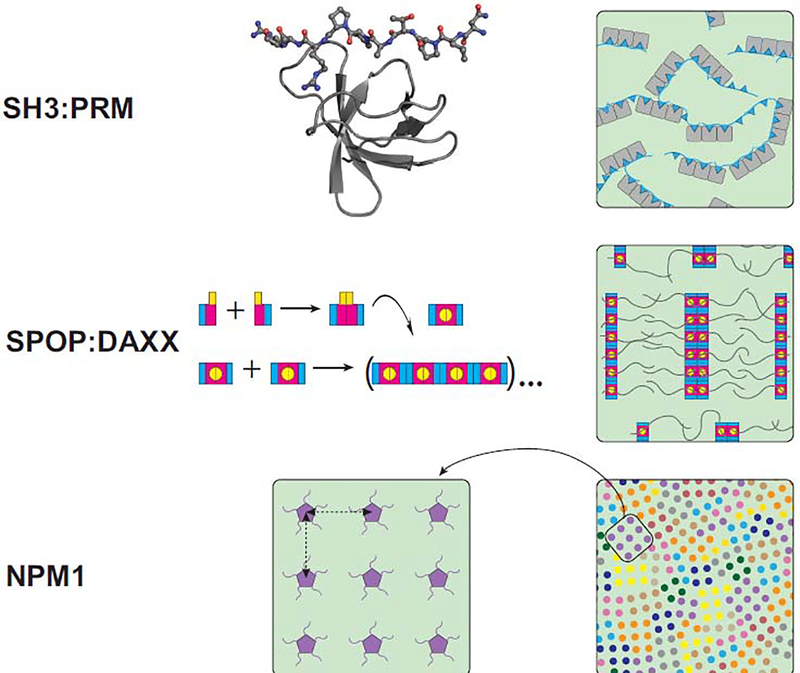
Evidence accumulated over the past decade provides support for liquid-liquid phase separation as the mechanism underlying the formation of biomolecular condensates, which include not only 'membraneless' organelles such as nucleoli and RNA granules, but additional assemblies involved in transcription, translation and signaling. Understanding the molecular mechanisms of condensate function requires knowledge of the structures of their constituents. Current knowledge suggests that structures formed via multivalent domain-motif interactions remain largely unchanged within condensates. Two different viewpoints exist regarding structures of disordered low-complexity domains within condensates; one argues that low-complexity domains remain largely disordered in condensates and their multivalency is encoded in short motifs called 'stickers', while the other argues that the sequences form cross-β structures resembling amyloid fibrils. We review these viewpoints and highlight outstanding questions that will inform structure-function relationships for biomolecular condensates.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures


References
-
- Franzmann TM, Jahnel M, Pozniakovsky A, Mahamid J, Holehouse AS, Nuske E, Richter D, Baumeister W, Grill SW, Pappu RV, et al. : Phase separation of a yeast prion protein promotes cellular fitness. Science 2018, 359. - PubMed
-
- Brangwynne CP, Eckmann CR, Courson DS, Rybarska A, Hoege C, Gharakhani J, Julicher F, Hyman AA: Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science 2009, 324:1729–1732. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
