

Telomerase increasing compound protects hippocampal neurons from amyloid beta toxicity by enhancing the expression of neurotrophins and plasticity related genes
- PMID: 31792359
- PMCID: PMC6889131
- DOI: 10.1038/s41598-019-54741-7
Telomerase increasing compound protects hippocampal neurons from amyloid beta toxicity by enhancing the expression of neurotrophins and plasticity related genes
Abstract
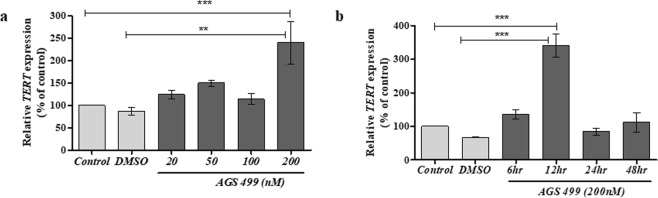
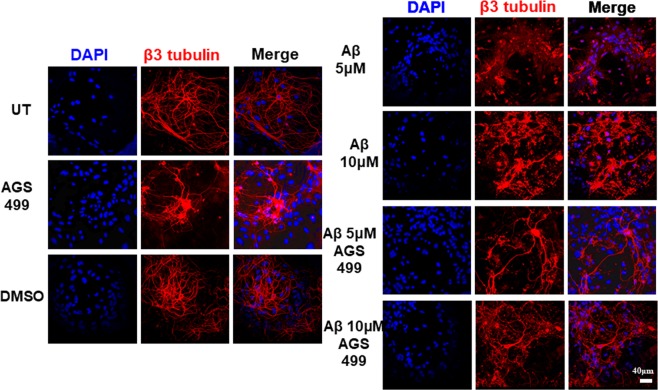
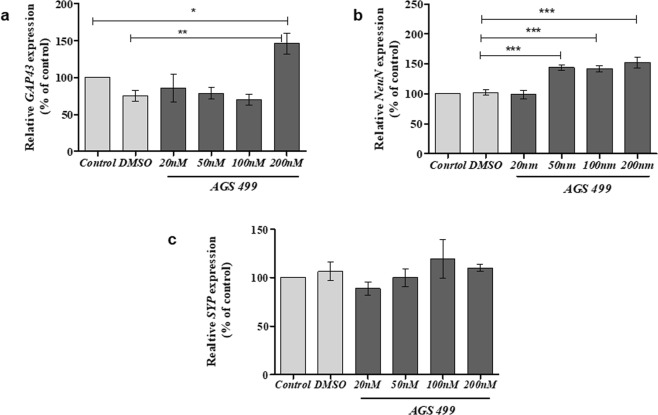
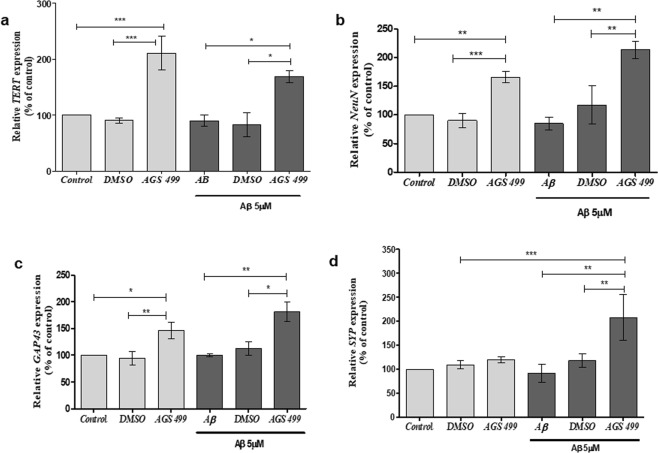
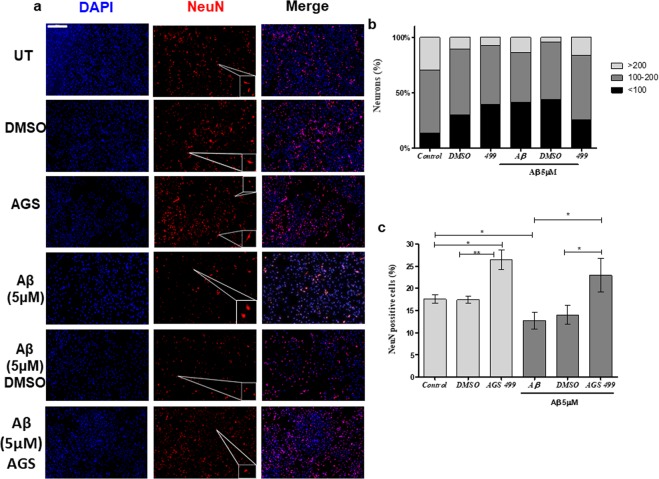
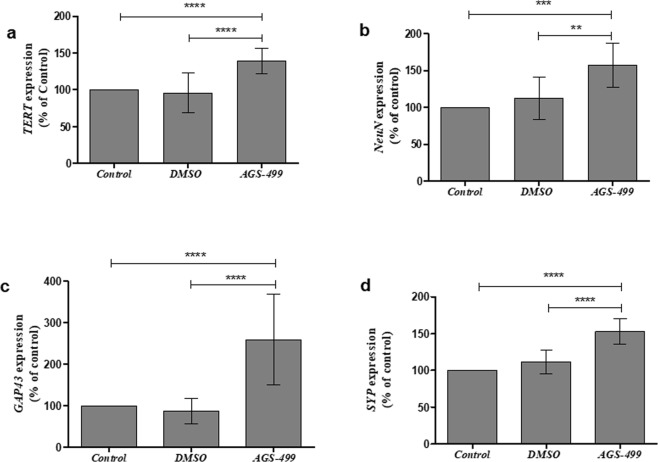
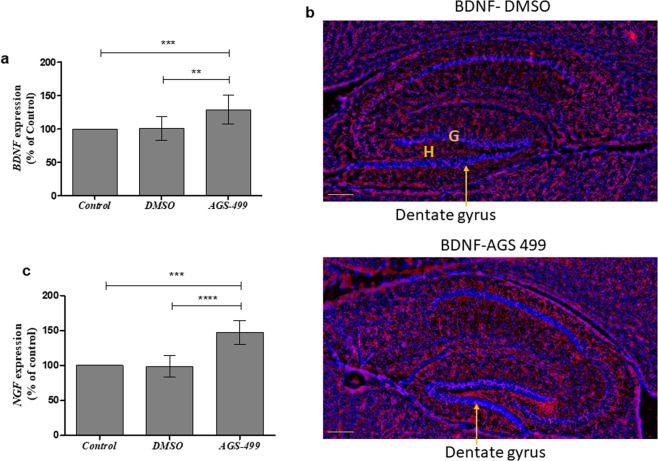
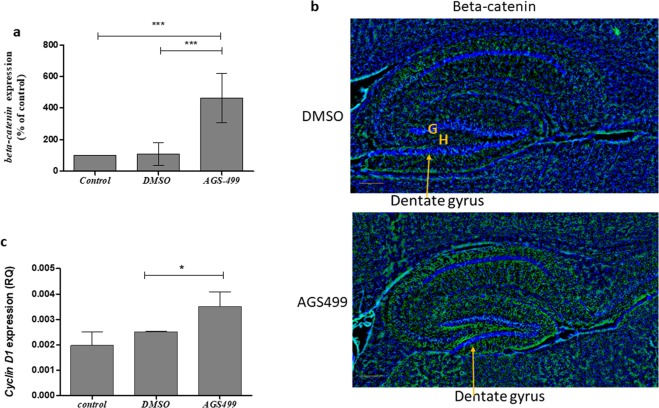
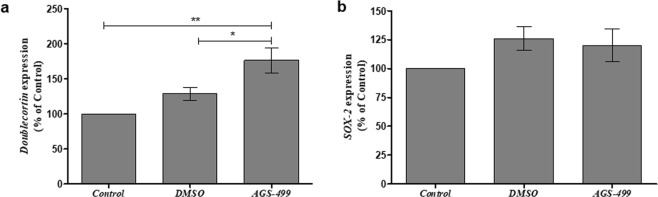
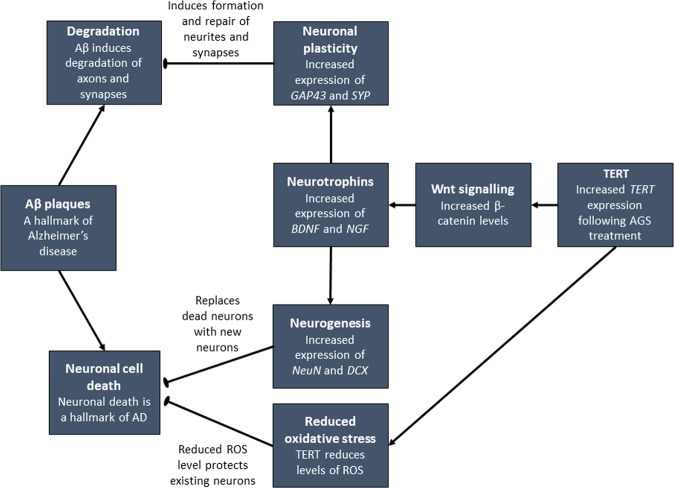
The telomerase reverse transcriptase protein, TERT, is expressed in the adult brain and its exogenic expression protects neurons from oxidative stress and from the cytotoxicity of amyloid beta (Aβ). We previously showed that telomerase increasing compounds (AGS) protected neurons from oxidative stress. Therefore, we suggest that increasing TERT by AGS may protect neurons from the Aβ-induced neurotoxicity by influencing genes and factors that participate in neuronal survival and plasticity. Here we used a primary hippocampal cell culture exposed to aggregated Aβ and hippocampi from adult mice. AGS treatment transiently increased TERT gene expression in hippocampal primary cell cultures in the presence or absence of Aβ and protected neurons from Aβ induced neuronal degradation. An increase in the expression of Growth associated protein 43 (GAP43), and Feminizing locus on X-3 genes (NeuN), in the presence or absence of Aβ, and Synaptophysin (SYP) in the presence of Aβ was observed. GAP43, NeuN, SYP, Neurotrophic factors (NGF, BDNF), beta-catenin and cyclin-D1 expression were increased in the hippocampus of AGS treated mice. This data suggests that increasing TERT by pharmaceutical compounds partially exerts its neuroprotective effect by enhancing the expression of neurotrophic factors and neuronal plasticity genes in a mechanism that involved Wnt/beta-catenin pathway.
Conflict of interest statement
E.P. filed a patent on the AGS compounds. N.B.R., V.R. and A.B. declare no potential conflict of interest. All authors declare no financial interest.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
