

LIN28B affects gene expression at the hypothalamic-pituitary axis and serum testosterone levels
- PMID: 31792362
- PMCID: PMC6889388
- DOI: 10.1038/s41598-019-54475-6
LIN28B affects gene expression at the hypothalamic-pituitary axis and serum testosterone levels
Abstract
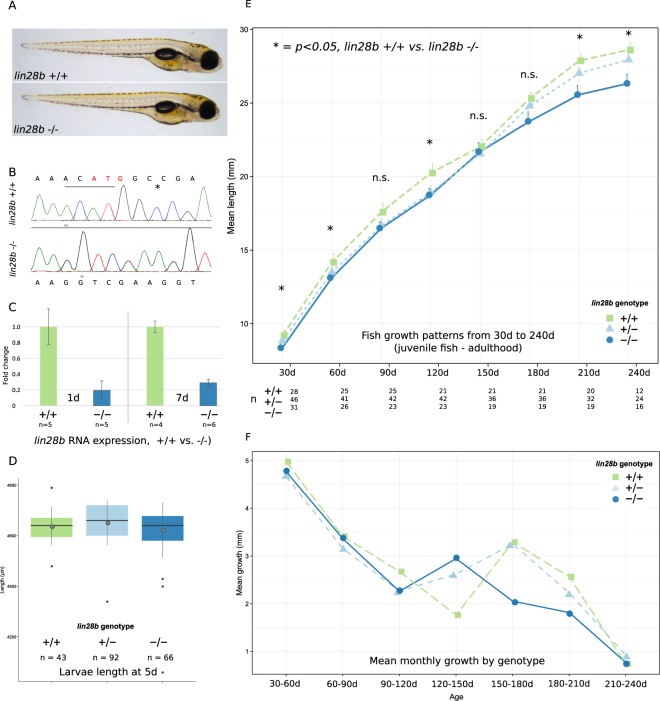
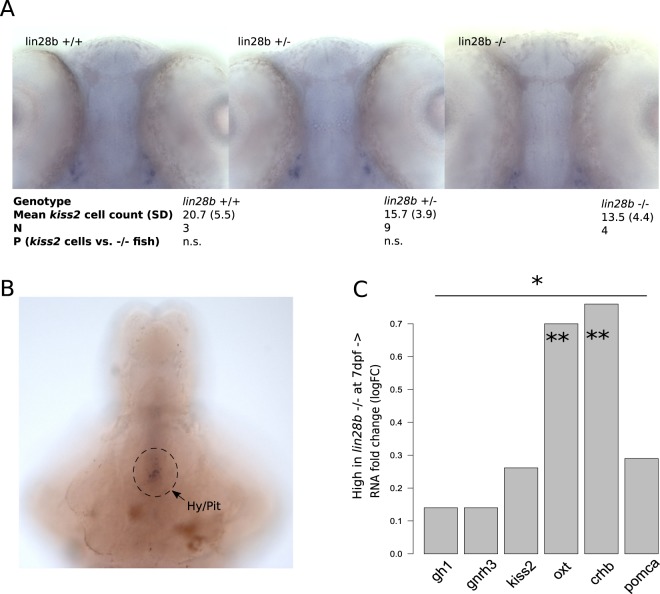
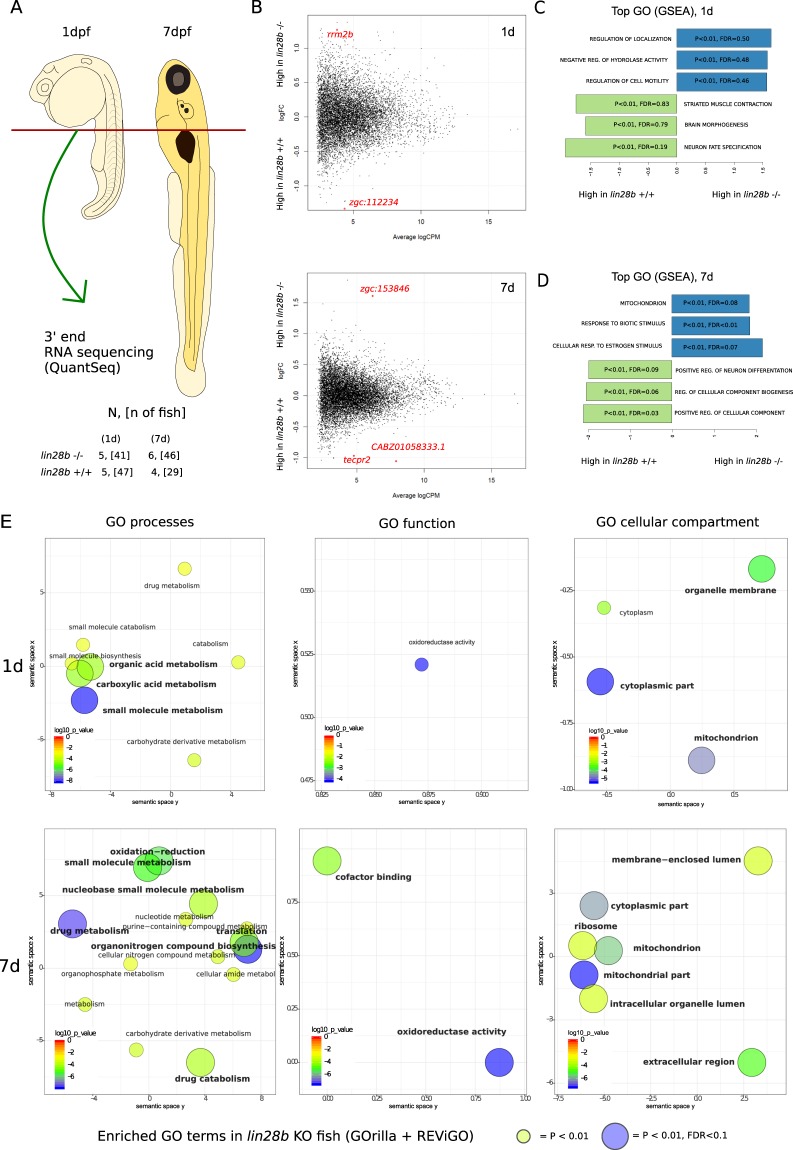
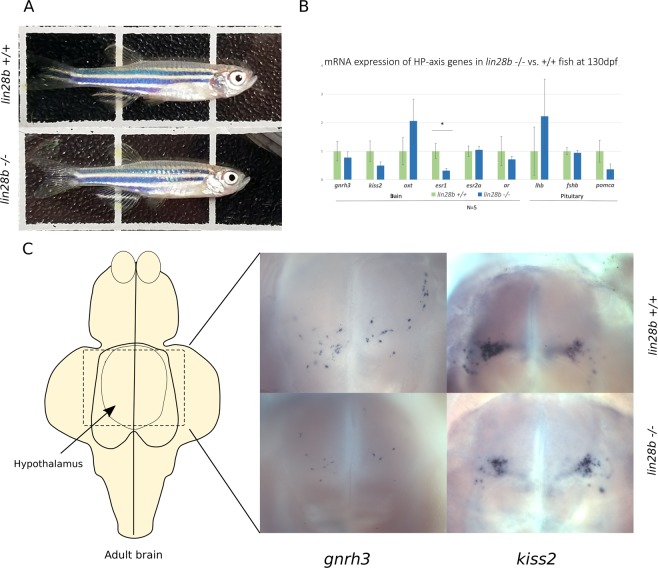
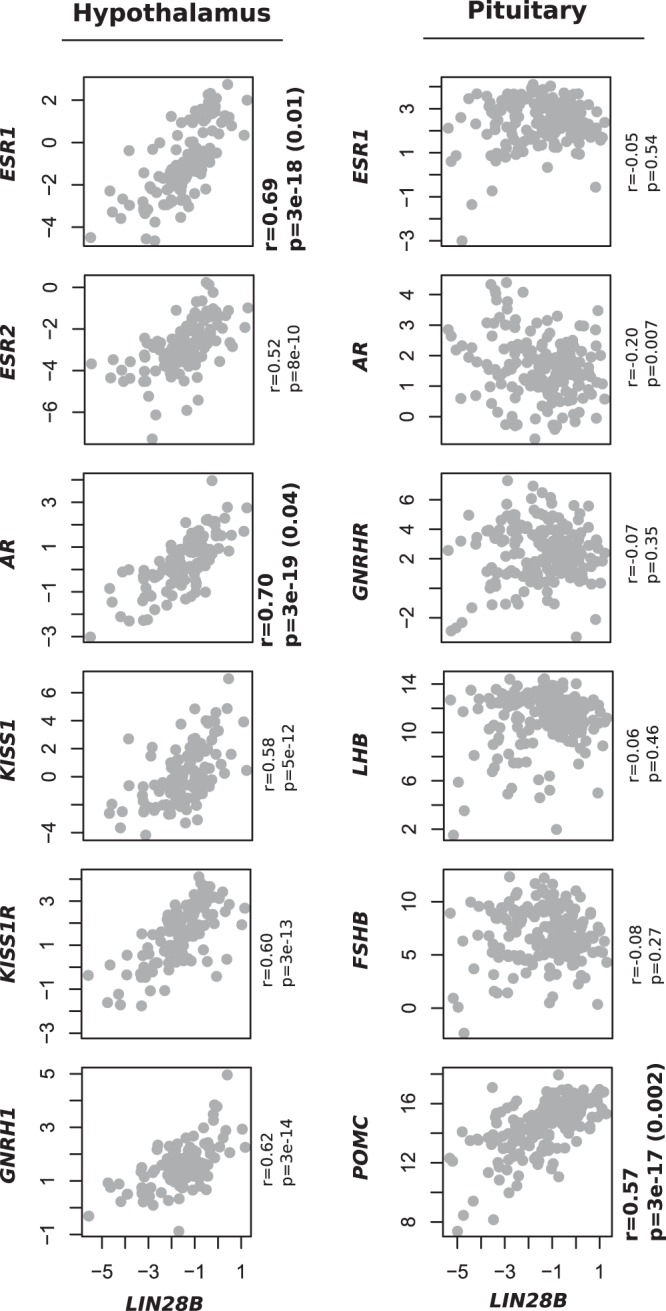
Genome-wide association studies (GWAS) have recurrently associated sequence variation nearby LIN28B with pubertal timing, growth and disease. However, the biology linking LIN28B with these traits is still poorly understood. With our study, we sought to elucidate the mechanisms behind the LIN28B associations, with a special focus on studying LIN28B function at the hypothalamic-pituitary (HP) axis that is ultimately responsible for pubertal onset. Using CRISPR-Cas9 technology, we first generated lin28b knockout (KO) zebrafish. Compared to controls, the lin28b KO fish showed both accelerated growth tempo, reduced adult size and increased expression of mitochondrial genes during larval development. Importantly, data from the knockout zebrafish models and adult humans imply that LIN28B expression has potential to affect gene expression in the HP axis. Specifically, our results suggest that LIN28B expression correlates positively with the expression of ESR1 in the hypothalamus and POMC in the pituitary. Moreover, we show how the pubertal timing advancing allele (T) for rs7759938 at the LIN28B locus associates with higher testosterone levels in the UK Biobank data. Overall, we provide novel evidence that LIN28B contributes to the regulation of sex hormone pathways, which might help explain why the gene associates with several distinct traits.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Day, F. R., Elks, C. E., Murray, A., Ong, K. K. & Perry, J. R. B. Puberty timing associated with diabetes, cardiovascular disease and also diverse health outcomes in men and women: the UK Biobank study. Sci. Rep. 5, 11208, 10.1038/srep11208, https://www.nature.com/articles/srep11208#supplementary-information (2015). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
