

A post-translational modification signature defines changes in soluble tau correlating with oligomerization in early stage Alzheimer's disease brain
- PMID: 31796124
- PMCID: PMC6892178
- DOI: 10.1186/s40478-019-0823-2
A post-translational modification signature defines changes in soluble tau correlating with oligomerization in early stage Alzheimer's disease brain
Abstract
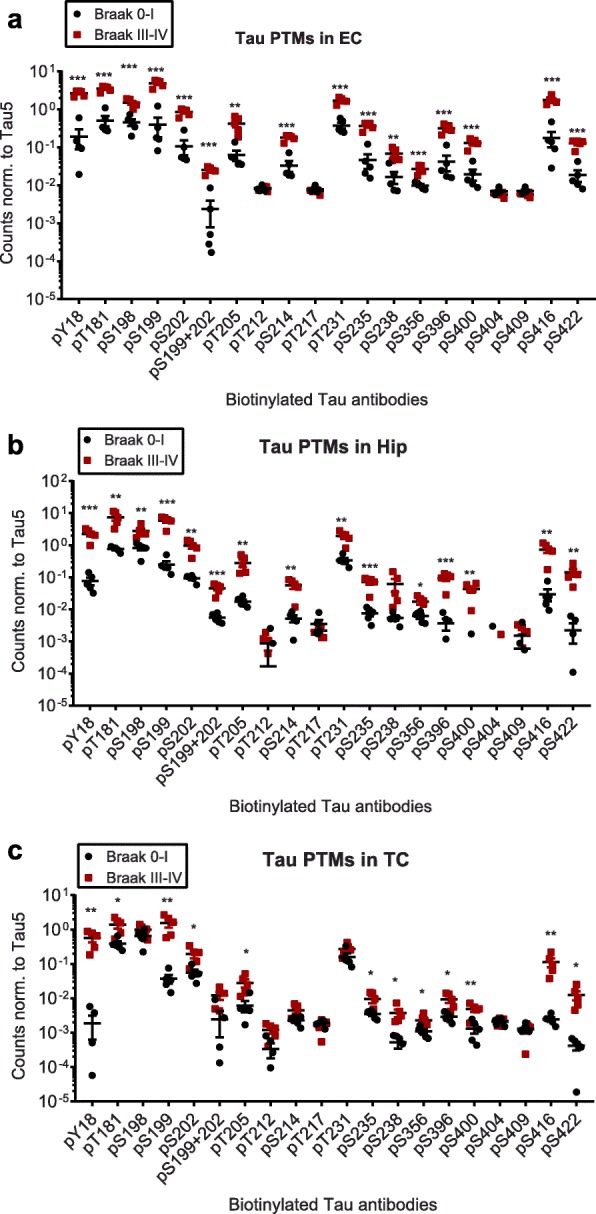
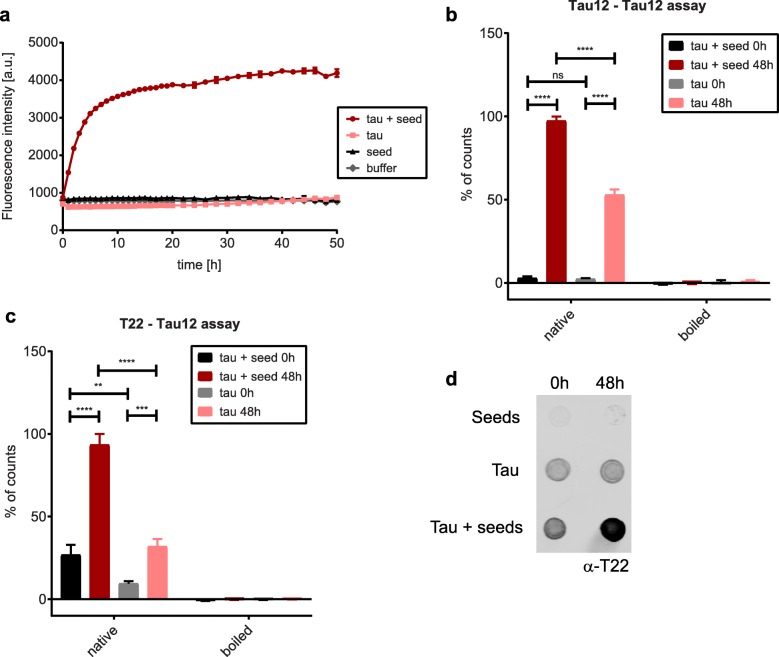
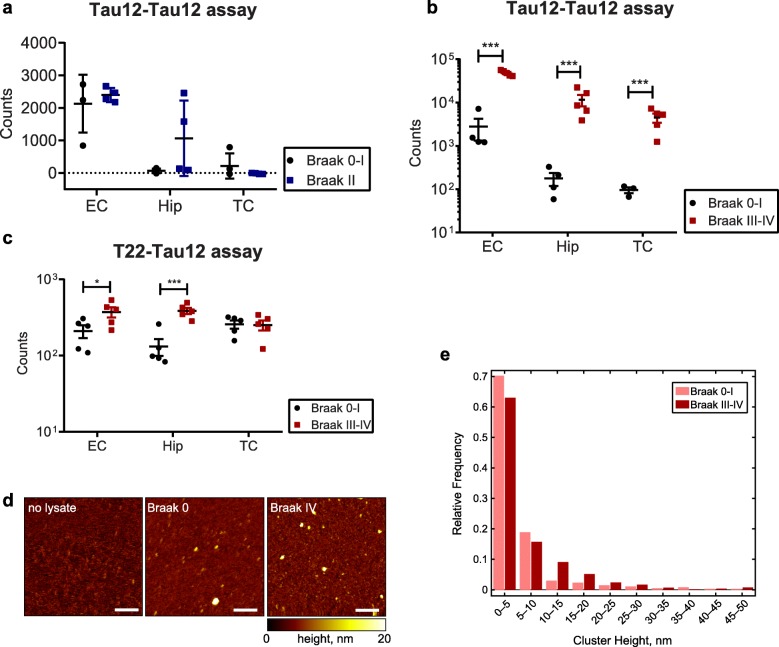
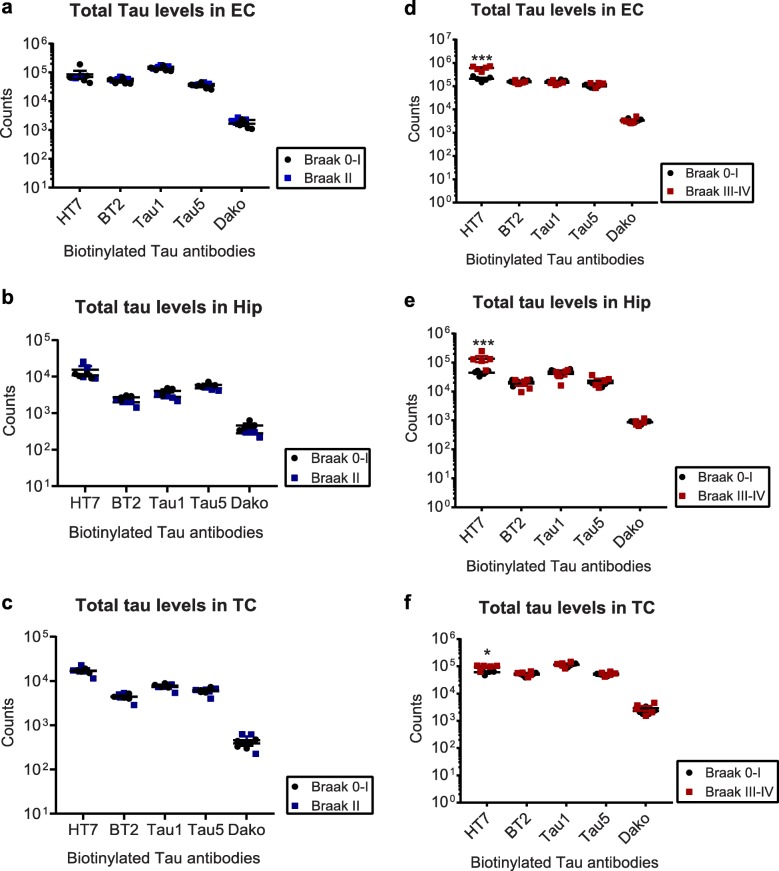
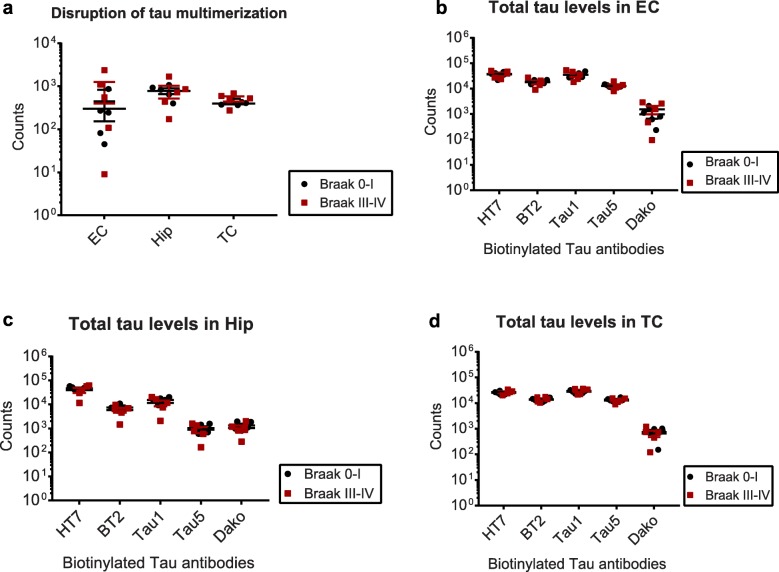
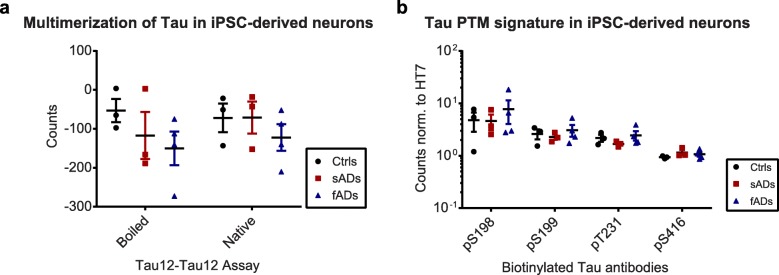
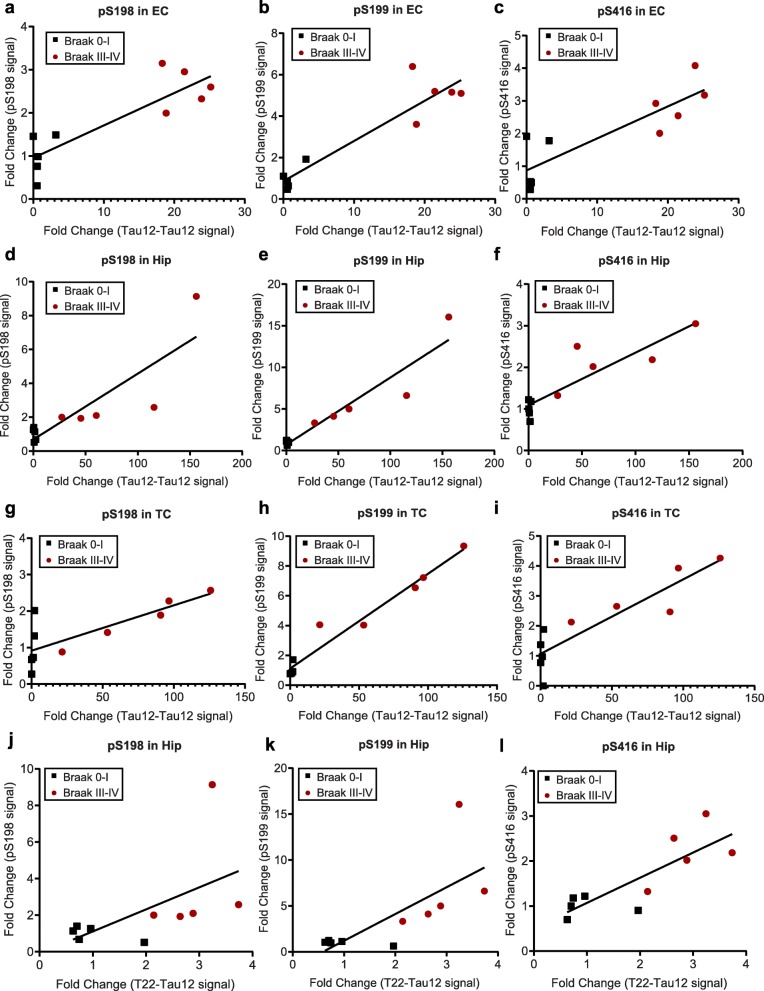
Tau is a microtubule-binding protein that can receive various post-translational modifications (PTMs) including phosphorylation, methylation, acetylation, glycosylation, nitration, sumoylation and truncation. Hyperphosphorylation of tau is linked to its aggregation and the formation of neurofibrillary tangles (NFTs), which are a hallmark of Alzheimer's disease (AD). While more than 70 phosphorylation sites have been detected previously on NFT tau, studies of oligomeric and detergent-soluble tau in human brains during the early stages of AD are lacking. Here we apply a comprehensive electrochemiluminescence ELISA assay to analyze twenty-five different PTM sites as well as tau oligomerization in control and sporadic AD brain. The samples were classified as Braak stages 0-I, II or III-IV, corresponding to the progression of microscopically detectable tau pathology throughout different brain regions. We found that soluble tau multimers are strongly increased at Braak stages III-IV in all brain regions under investigation, including the temporal cortex, which does not contain NFTs or misfolded oligomers at this stage of pathology. We additionally identified five phosphorylation sites that are specifically and consistently increased across the entorhinal cortex, hippocampus and temporal cortex in the same donors. Three of these sites correlate with tau multimerization in all three brain regions, but do not overlap with the epitopes of phospho-sensitive antibodies commonly used for the immunohistochemical detection of NFTs. Our results thus suggest that soluble multimers are characterized by a small set of specific phosphorylation events that differ from those dominating in mature NFTs. These findings shed light on early PTM changes of tau during AD pathogenesis in human brains.
Keywords: Alzheimer’s disease; Posttranslational modifications; Tau; Tau oligomerization.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
-
- Alzheimer A (1907) Über eine eigenartige Erkrankung der Hirnrinde. Allgemeine Zeitschrift fur Psychiatrie und Psychisch-gerichtliche Medizin 64: 146–148
-
- Ince PG, McArthur FK, Bjertness E, Torvik A, Candy JM, Edwardson JA (1995) Neuropathological diagnoses in elderly patients in Oslo: Alzheimer's disease, Lewy body disease, vascular lesions. Dementia 6: 162–168 - PubMed
-
- Dickson DW (1997) Neuropathological diagnosis of Alzheimer's disease: a perspective from longitudinal clinicopathological studies. Neurobiol Aging 18: S21–26 - PubMed
-
- Braak H, Braak E (1998) Evolution of neuronal changes in the course of Alzheimer's disease. J Neural Transm Suppl 53: 127–140 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
