

Biology of the Caenorhabditis elegans Germline Stem Cell System
- PMID: 31796552
- PMCID: PMC6893382
- DOI: 10.1534/genetics.119.300238
Biology of the Caenorhabditis elegans Germline Stem Cell System
Abstract
Stem cell systems regulate tissue development and maintenance. The germline stem cell system is essential for animal reproduction, controlling both the timing and number of progeny through its influence on gamete production. In this review, we first draw general comparisons to stem cell systems in other organisms, and then present our current understanding of the germline stem cell system in Caenorhabditis elegans In contrast to stereotypic somatic development and cell number stasis of adult somatic cells in C. elegans, the germline stem cell system has a variable division pattern, and the system differs between larval development, early adult peak reproduction and age-related decline. We discuss the cell and developmental biology of the stem cell system and the Notch regulated genetic network that controls the key decision between the stem cell fate and meiotic development, as it occurs under optimal laboratory conditions in adult and larval stages. We then discuss alterations of the stem cell system in response to environmental perturbations and aging. A recurring distinction is between processes that control stem cell fate and those that control cell cycle regulation. C. elegans is a powerful model for understanding germline stem cells and stem cell biology.
Keywords: Caenorhabditis elegans; Notch; WormBook; network; niche; physiology; stem cell.
Copyright © 2019 by the Genetics Society of America.
Figures
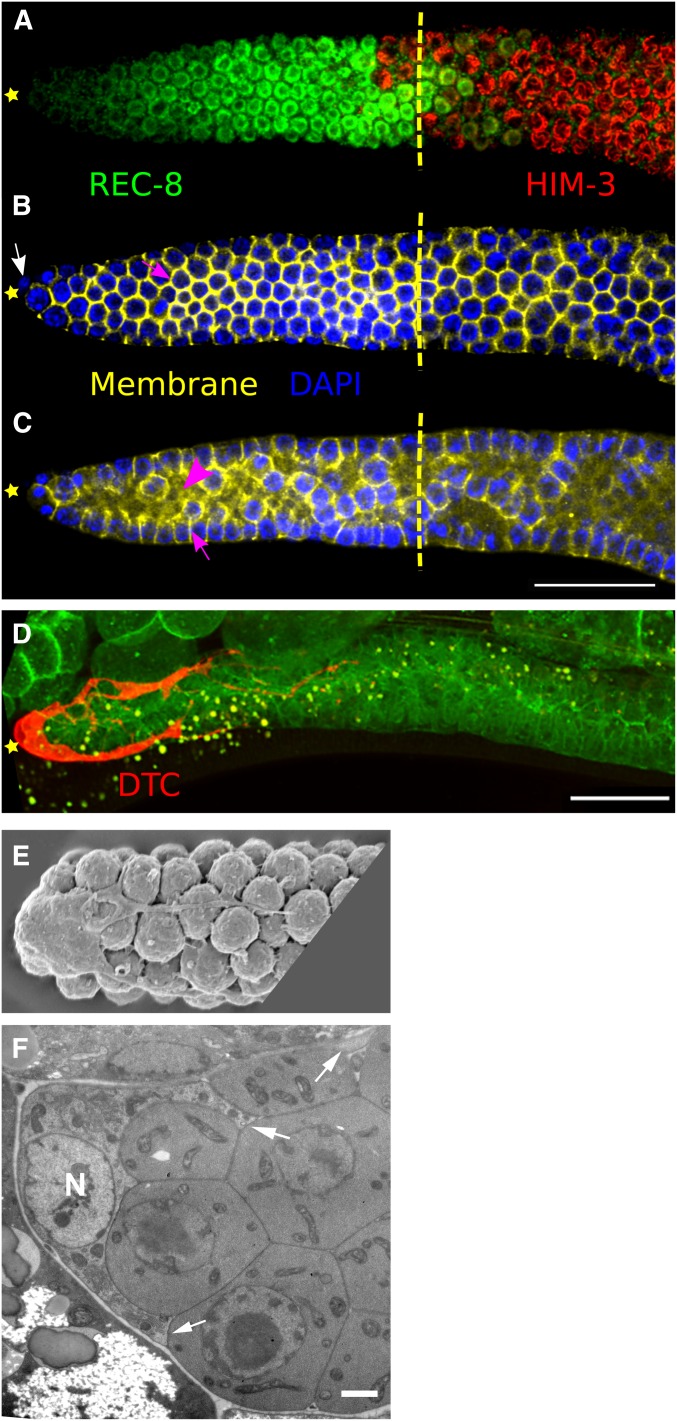
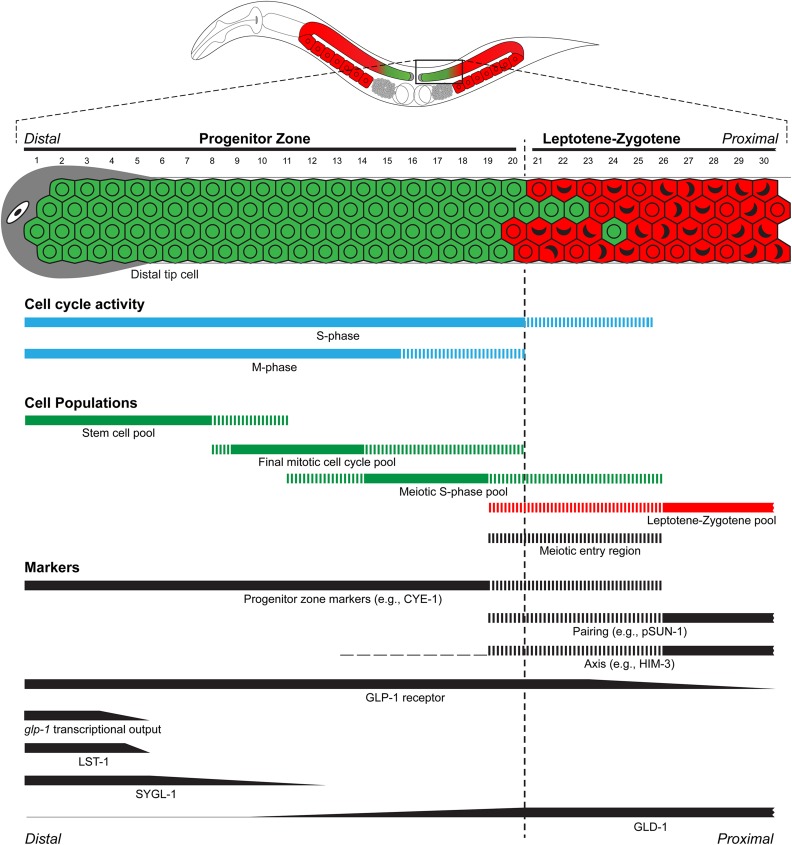
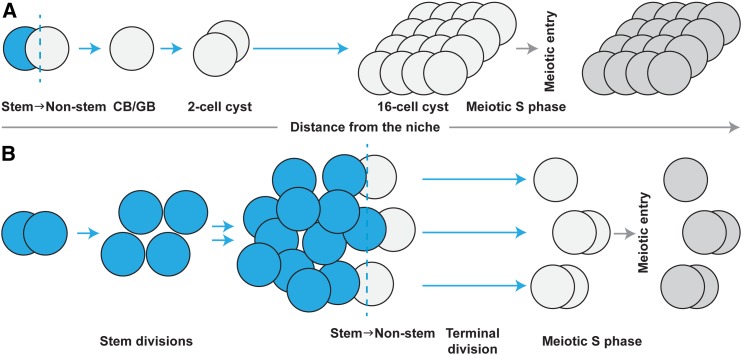
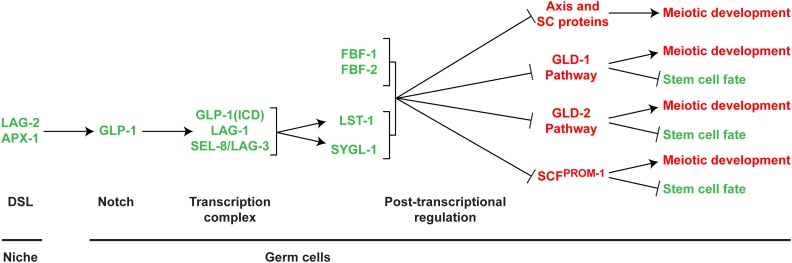
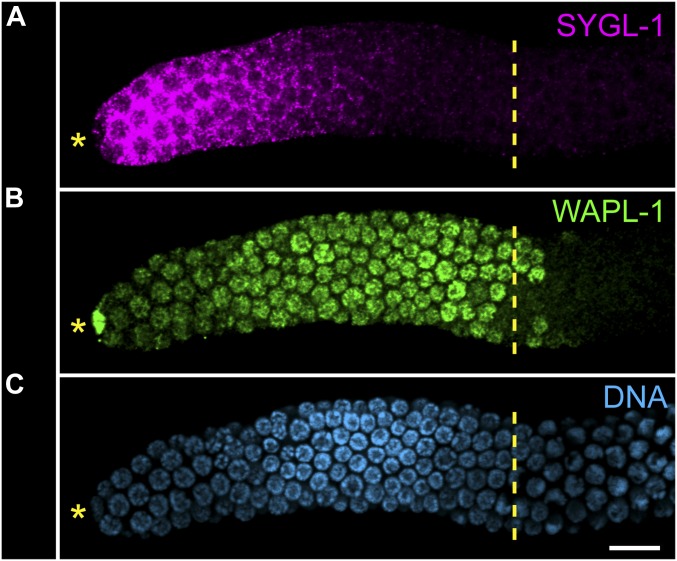
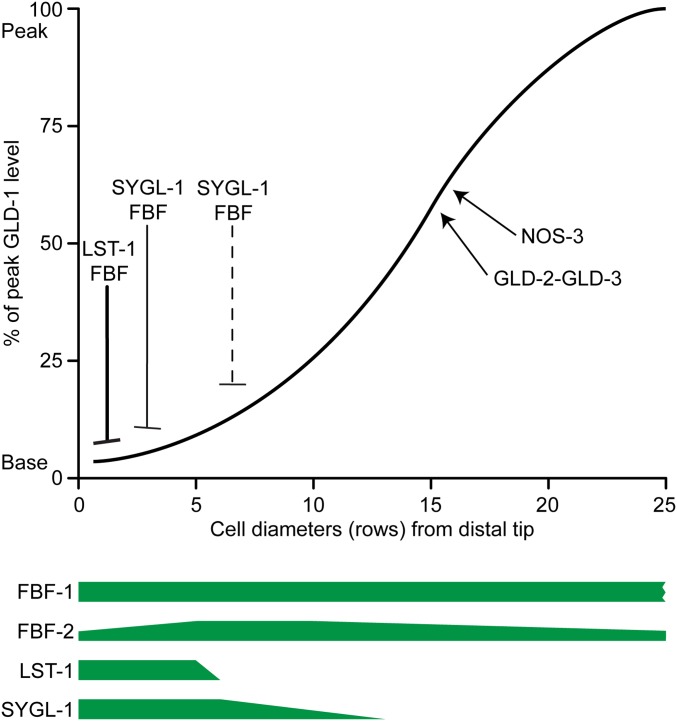
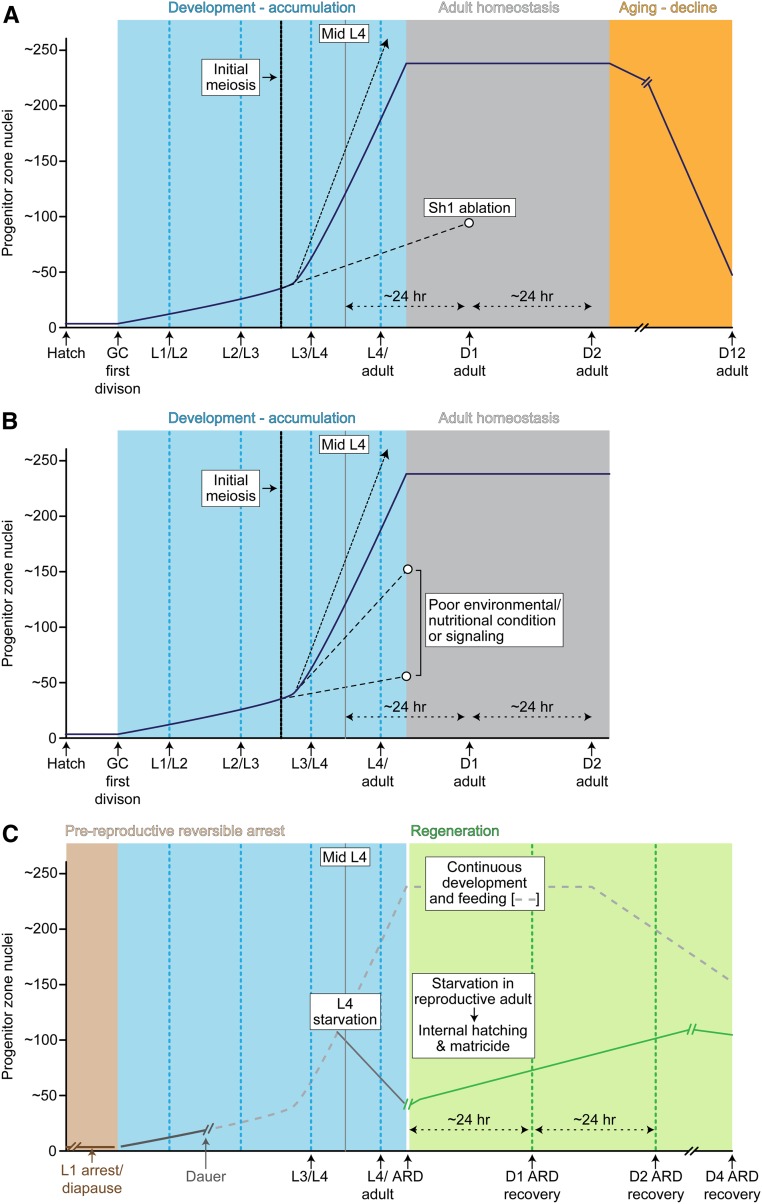
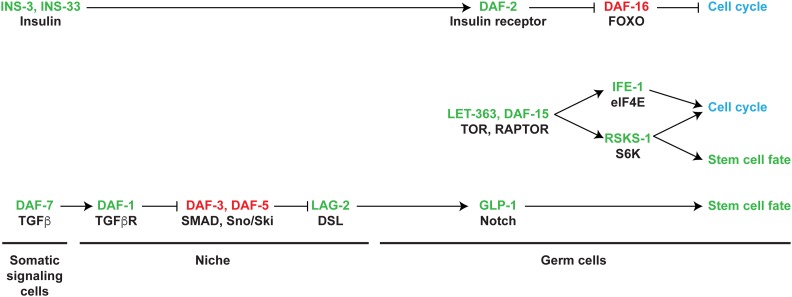
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
