

The coincidence of ecological opportunity with hybridization explains rapid adaptive radiation in Lake Mweru cichlid fishes
- PMID: 31796733
- PMCID: PMC6890737
- DOI: 10.1038/s41467-019-13278-z
The coincidence of ecological opportunity with hybridization explains rapid adaptive radiation in Lake Mweru cichlid fishes
Abstract
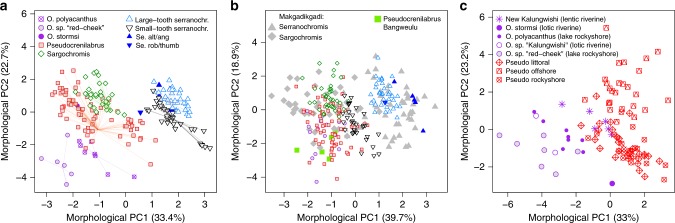
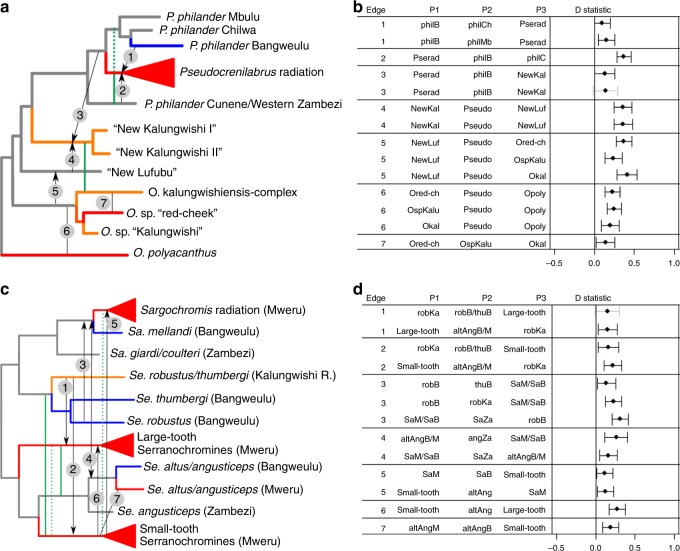
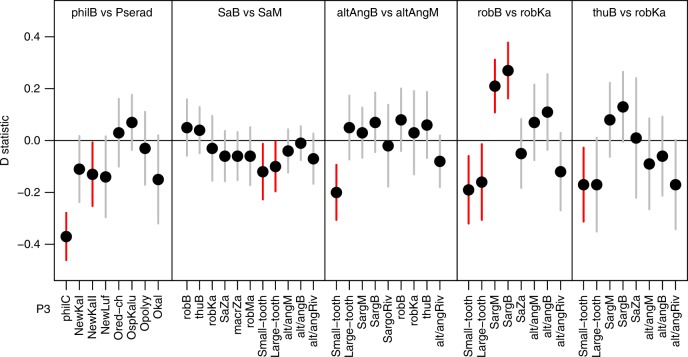
The process of adaptive radiation was classically hypothesized to require isolation of a lineage from its source (no gene flow) and from related species (no competition). Alternatively, hybridization between species may generate genetic variation that facilitates adaptive radiation. Here we study haplochromine cichlid assemblages in two African Great Lakes to test these hypotheses. Greater biotic isolation (fewer lineages) predicts fewer constraints by competition and hence more ecological opportunity in Lake Bangweulu, whereas opportunity for hybridization predicts increased genetic potential in Lake Mweru. In Lake Bangweulu, we find no evidence for hybridization but also no adaptive radiation. We show that the Bangweulu lineages also colonized Lake Mweru, where they hybridized with Congolese lineages and then underwent multiple adaptive radiations that are strikingly complementary in ecology and morphology. Our data suggest that the presence of several related lineages does not necessarily prevent adaptive radiation, although it constrains the trajectories of morphological diversification. It might instead facilitate adaptive radiation when hybridization generates genetic variation, without which radiation may start much later, progress more slowly or never occur.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Ecological opportunity and sexual selection together predict adaptive radiation.Nature. 2012 Jul 19;487(7407):366-9. doi: 10.1038/nature11144. Nature. 2012. PMID: 22722840
-
Relaxed trait covariance in interspecific cichlid hybrids predicts morphological diversity in adaptive radiations.J Evol Biol. 2014 Jan;27(1):11-24. doi: 10.1111/jeb.12283. Epub 2013 Dec 16. J Evol Biol. 2014. PMID: 24330234
-
Ancient hybridization fuels rapid cichlid fish adaptive radiations.Nat Commun. 2017 Feb 10;8:14363. doi: 10.1038/ncomms14363. Nat Commun. 2017. PMID: 28186104 Free PMC article.
-
African cichlid fish: a model system in adaptive radiation research.Proc Biol Sci. 2006 Aug 22;273(1597):1987-98. doi: 10.1098/rspb.2006.3539. Proc Biol Sci. 2006. PMID: 16846905 Free PMC article. Review.
-
Local variation and parallel evolution: morphological and genetic diversity across a species complex of neotropical crater lake cichlid fishes.Philos Trans R Soc Lond B Biol Sci. 2010 Jun 12;365(1547):1763-82. doi: 10.1098/rstb.2009.0271. Philos Trans R Soc Lond B Biol Sci. 2010. PMID: 20439280 Free PMC article. Review.
Cited by
-
Detection and quantification of introgression using Bayesian inference based on conjugate priors.Bioinformatics. 2024 Nov 1;40(11):btae642. doi: 10.1093/bioinformatics/btae642. Bioinformatics. 2024. PMID: 39460951 Free PMC article.
-
Rapid generation of ecologically relevant behavioral novelty in experimental cichlid hybrids.Ecol Evol. 2020 Jun 16;10(14):7445-7462. doi: 10.1002/ece3.6471. eCollection 2020 Jul. Ecol Evol. 2020. PMID: 32760540 Free PMC article.
-
Ecological outcomes of hybridization vary extensively in Catostomus fishes.Evolution. 2022 Nov;76(11):2697-2711. doi: 10.1111/evo.14624. Epub 2022 Sep 27. Evolution. 2022. PMID: 36097356 Free PMC article.
-
A vertebrate adaptive radiation is assembled from an ancient and disjunct spatiotemporal landscape.Proc Natl Acad Sci U S A. 2021 May 18;118(20):e2011811118. doi: 10.1073/pnas.2011811118. Proc Natl Acad Sci U S A. 2021. PMID: 33990463 Free PMC article.
-
Evolutionary dynamics of pre- and postzygotic reproductive isolation in cichlid fishes.Philos Trans R Soc Lond B Biol Sci. 2020 Aug 31;375(1806):20190535. doi: 10.1098/rstb.2019.0535. Epub 2020 Jul 13. Philos Trans R Soc Lond B Biol Sci. 2020. PMID: 32654645 Free PMC article. Review.
References
-
- Schluter, D. The Ecology Of Adaptive Radiation (Oxford University Press Inc., 2000).
-
- Simpson, G. G. Evolution and Geography; An Essay On Historical Biogeography, With Special Reference To Mammals. (Oregon State System of Higher Education, 1953).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
