

Orthogonal regulation of DNA nanostructure self-assembly and disassembly using antibodies
- PMID: 31796740
- PMCID: PMC6890650
- DOI: 10.1038/s41467-019-13104-6
Orthogonal regulation of DNA nanostructure self-assembly and disassembly using antibodies
Erratum in
-
Publisher Correction: Orthogonal regulation of DNA nanostructure self-assembly and disassembly using antibodies.Nat Commun. 2020 Jan 13;11(1):327. doi: 10.1038/s41467-019-13971-z. Nat Commun. 2020. PMID: 31932618 Free PMC article.
Abstract
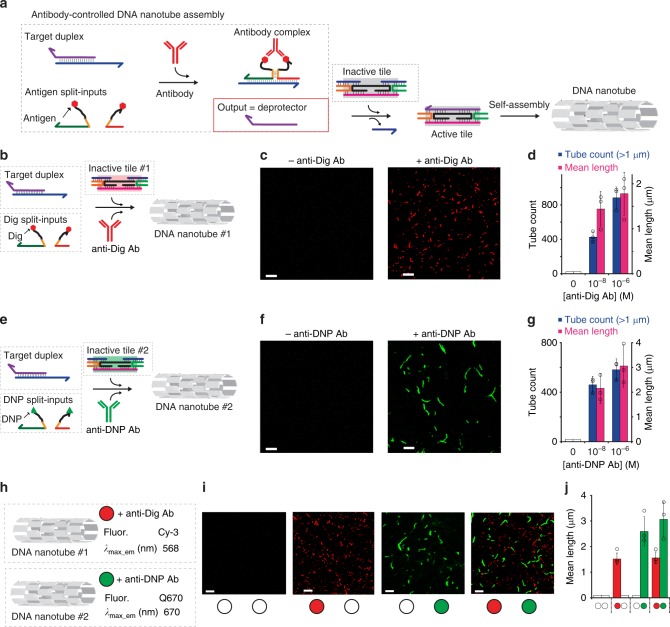
Here we report a rational strategy to orthogonally control assembly and disassembly of DNA-based nanostructures using specific IgG antibodies as molecular inputs. We first demonstrate that the binding of a specific antibody to a pair of antigen-conjugated split DNA input-strands induces their co-localization and reconstitution into a functional unit that is able to initiate a toehold strand displacement reaction. The effect is rapid and specific and can be extended to different antibodies with the expedient of changing the recognition elements attached to the two split DNA input-strands. Such an antibody-regulated DNA-based circuit has then been employed to control the assembly and disassembly of DNA tubular structures using specific antibodies as inputs. For example, we demonstrate that we can induce self-assembly and disassembly of two distinct DNA tubular structures by using DNA circuits controlled by two different IgG antibodies (anti-Dig and anti-DNP antibodies) in the same solution in an orthogonal way.
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Light-Controlled, Toehold-Mediated Logic Circuit for Assembly of DNA Tiles.ACS Appl Mater Interfaces. 2020 Feb 5;12(5):6336-6342. doi: 10.1021/acsami.9b21778. Epub 2020 Jan 23. ACS Appl Mater Interfaces. 2020. PMID: 31918539
-
Remote Electronic Control of DNA-Based Reactions and Nanostructure Assembly.Nano Lett. 2018 May 9;18(5):2918-2923. doi: 10.1021/acs.nanolett.8b00179. Epub 2018 Apr 5. Nano Lett. 2018. PMID: 29608313
-
pH-Driven Reversible Self-Assembly of Micron-Scale DNA Scaffolds.Nano Lett. 2017 Dec 13;17(12):7283-7288. doi: 10.1021/acs.nanolett.7b02787. Epub 2017 Nov 28. Nano Lett. 2017. PMID: 29182337
-
Sequence-specific recognition of DNA nanostructures.Methods. 2014 May 15;67(2):123-33. doi: 10.1016/j.ymeth.2014.02.028. Epub 2014 Feb 28. Methods. 2014. PMID: 24583116 Review.
-
Dynamic DNA nanotechnology using strand-displacement reactions.Nat Chem. 2011 Feb;3(2):103-13. doi: 10.1038/nchem.957. Nat Chem. 2011. PMID: 21258382 Review.
Cited by
-
Programmable Cell-Free Transcriptional Switches for Antibody Detection.J Am Chem Soc. 2022 Apr 6;144(13):5820-5826. doi: 10.1021/jacs.1c11706. Epub 2022 Mar 22. J Am Chem Soc. 2022. PMID: 35316049 Free PMC article.
-
Programmable, Multiplexed DNA Circuits Supporting Clinically Relevant, Electrochemical Antibody Detection.ACS Sens. 2021 Jun 25;6(6):2442-2448. doi: 10.1021/acssensors.1c00790. Epub 2021 Jun 15. ACS Sens. 2021. PMID: 34129321 Free PMC article.
-
Switchable supracolloidal 3D DNA origami nanotubes mediated through fuel/antifuel reactions.Nanoscale. 2020 Aug 20;12(32):16995-17004. doi: 10.1039/d0nr04209a. Nanoscale. 2020. PMID: 32780076 Free PMC article.
-
Encoding, Decoding, and Rendering Information in DNA Nanoswitch Libraries.ACS Synth Biol. 2023 Apr 21;12(4):978-983. doi: 10.1021/acssynbio.2c00649. Epub 2022 Dec 21. ACS Synth Biol. 2023. PMID: 36541933 Free PMC article.
-
Dissipative DNA nanotechnology.Nat Chem. 2022 Jun;14(6):600-613. doi: 10.1038/s41557-022-00957-6. Epub 2022 Jun 6. Nat Chem. 2022. PMID: 35668213 Review.
References
-
- Tan EM. Antinuclear antibodies: diagnostic markers for autoimmune diseases and probes for cell biology. Adv. Immunol. 1989;44:93–151. - PubMed
-
- Gubala V, Harris LF, Ricco AJ, Tan MX, Williams DE. Point of care diagnostics: status and future. Anal. Chem. 2012;84:487–515. - PubMed
-
- Coulie PG, Van den Eynde BJ, van der Bruggen P, Boon T. Tumour antigens recognized by T lymphocytes: at the core of cancer immunotherapy. Nat. Rev. Cancer. 2014;14:135–146. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
