

Elevated CO2 regulates the Wnt signaling pathway in mammals, Drosophila melanogaster and Caenorhabditis elegans
- PMID: 31796806
- PMCID: PMC6890671
- DOI: 10.1038/s41598-019-54683-0
Elevated CO2 regulates the Wnt signaling pathway in mammals, Drosophila melanogaster and Caenorhabditis elegans
Abstract
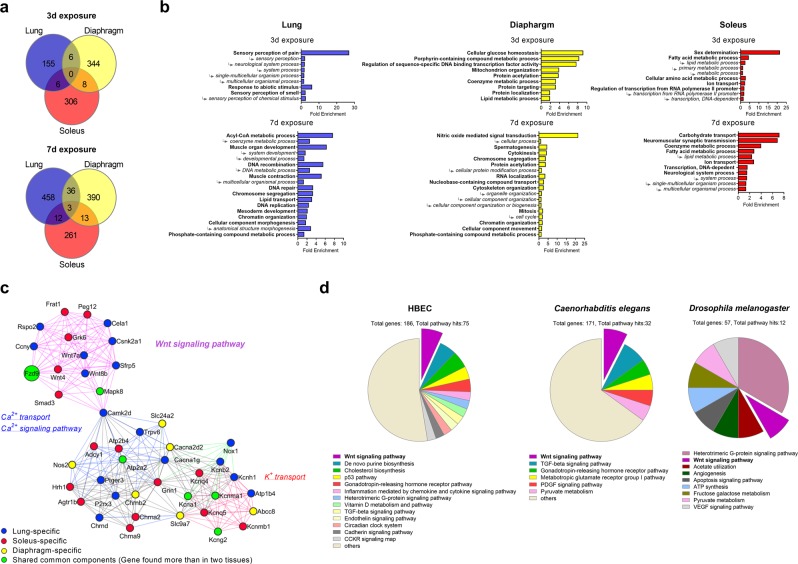
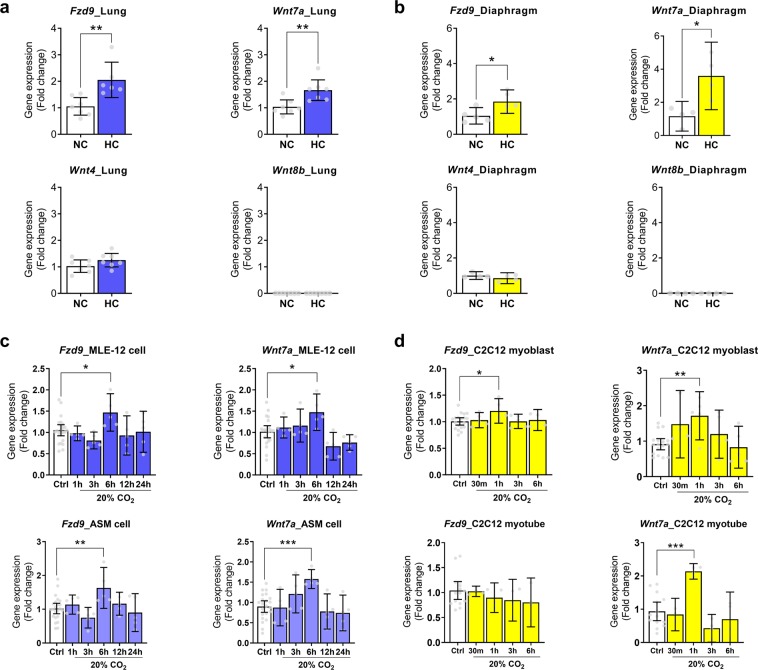
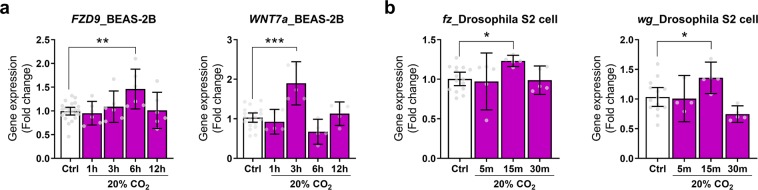
Carbon dioxide (CO2) is sensed by cells and can trigger signals to modify gene expression in different tissues leading to changes in organismal functions. Despite accumulating evidence that several pathways in various organisms are responsive to CO2 elevation (hypercapnia), it has yet to be elucidated how hypercapnia activates genes and signaling pathways, or whether they interact, are integrated, or are conserved across species. Here, we performed a large-scale transcriptomic study to explore the interaction/integration/conservation of hypercapnia-induced genomic responses in mammals (mice and humans) as well as invertebrates (Caenorhabditis elegans and Drosophila melanogaster). We found that hypercapnia activated genes that regulate Wnt signaling in mouse lungs and skeletal muscles in vivo and in several cell lines of different tissue origin. Hypercapnia-responsive Wnt pathway homologues were similarly observed in secondary analysis of available transcriptomic datasets of hypercapnia in a human bronchial cell line, flies and nematodes. Our data suggest the evolutionarily conserved role of high CO2 in regulating Wnt pathway genes.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
