

Mix-and-inject XFEL crystallography reveals gated conformational dynamics during enzyme catalysis
- PMID: 31801874
- PMCID: PMC6926069
- DOI: 10.1073/pnas.1901864116
Mix-and-inject XFEL crystallography reveals gated conformational dynamics during enzyme catalysis
Abstract
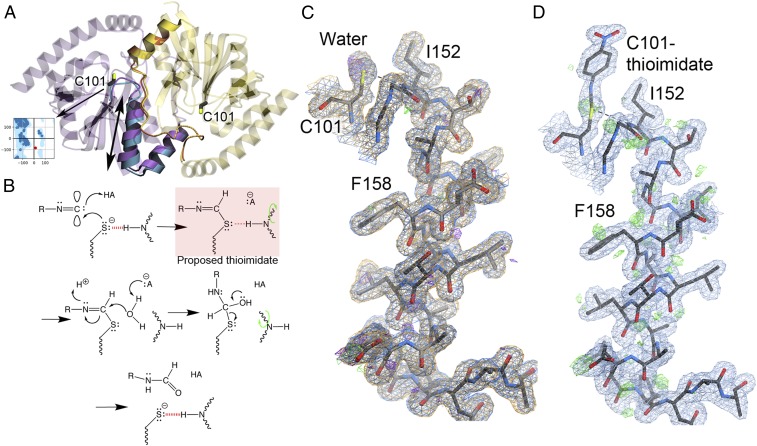
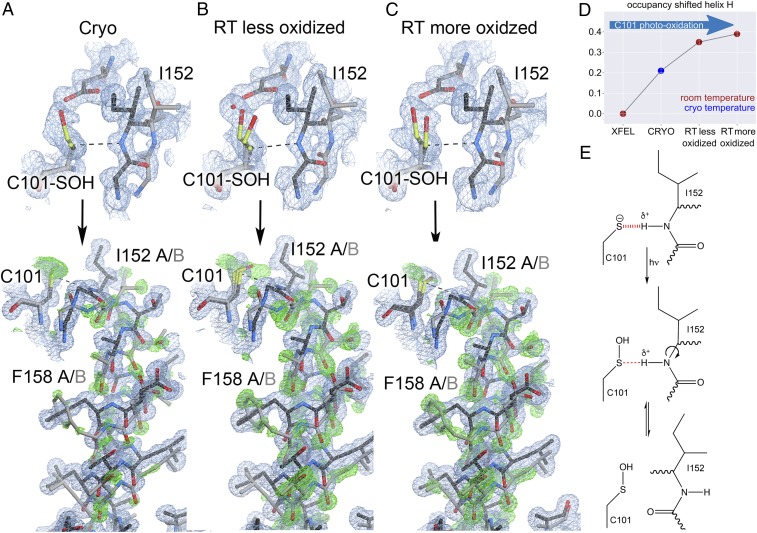
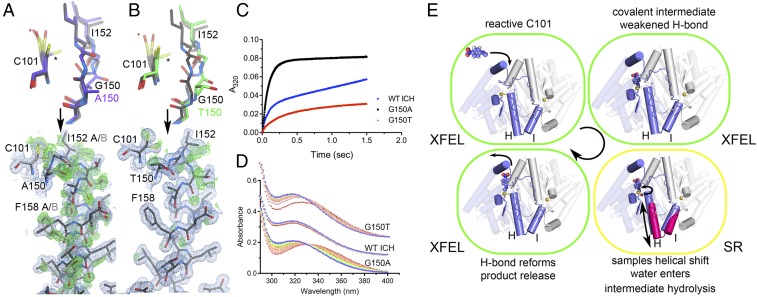
How changes in enzyme structure and dynamics facilitate passage along the reaction coordinate is a fundamental unanswered question. Here, we use time-resolved mix-and-inject serial crystallography (MISC) at an X-ray free electron laser (XFEL), ambient-temperature X-ray crystallography, computer simulations, and enzyme kinetics to characterize how covalent catalysis modulates isocyanide hydratase (ICH) conformational dynamics throughout its catalytic cycle. We visualize this previously hypothetical reaction mechanism, directly observing formation of a thioimidate covalent intermediate in ICH microcrystals during catalysis. ICH exhibits a concerted helical displacement upon active-site cysteine modification that is gated by changes in hydrogen bond strength between the cysteine thiolate and the backbone amide of the highly strained Ile152 residue. These catalysis-activated motions permit water entry into the ICH active site for intermediate hydrolysis. Mutations at a Gly residue (Gly150) that modulate helical mobility reduce ICH catalytic turnover and alter its pre-steady-state kinetic behavior, establishing that helical mobility is important for ICH catalytic efficiency. These results demonstrate that MISC can capture otherwise elusive aspects of enzyme mechanism and dynamics in microcrystalline samples, resolving long-standing questions about the connection between nonequilibrium protein motions and enzyme catalysis.
Keywords: X-ray crystallography; XFEL; cysteine modification; enzyme conformational dynamics; radiation damage.
Conflict of interest statement
The authors declare no competing interest.
Figures

References
-
- Henzler-Wildman K. A., et al. , Intrinsic motions along an enzymatic reaction trajectory. Nature 450, 838–844 (2007). - PubMed
-
- Kim T. H., et al. , The role of dimer asymmetry and protomer dynamics in enzyme catalysis. Science 355, eaag2355 (2017). - PubMed
-
- Hammes G. G., Multiple conformational changes in enzyme catalysis. Biochemistry 41, 8221–8228 (2002). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
